Introduction
Organophosphates (OP) are a bunch of pesticides that are widely used for a variety of agricultural, medical, and industrial applications. These compounds inhibit acetylcholine-esterase (AChE) enzyme activity, which causes the accumulation of acetylcholine (ACh) in the synaptic cleft that causes hypercholinergic symptoms. Poisoning by OPs is one of the global health problems and are responsible for the third most common cause of poisoning in Iran [1–3]. Diazinon (DZN) is one of the most important OP insecticides that is used extensively for various health-related and agricultural purposes pesticides in Iran [4]. Its residues in foods, water, vegetables and soil have direct effects on health of humans and animals. DZN is a highly toxic OP pesticide that oxidatively bioactivates to diazoxon by the liver microsomal enzymes system [3,5]. DZN causes some hematological, biochemical and histological alterations in rat, rabbits and mice. Its exposure may also cause alterations many systems such as the immune system, urinary system, reproductive system, pancreas and liver [2,6]. Furthermore, DZN produces oxidative stress through the formation of reactive oxygen species (ROS) and alterations in antioxidants status in mammals and other organisms [2,7,8].
Antioxidants constitute the primary defense system that limits the toxicity associated with ROS. DZN is a lipophilic compound with a long half-life that binds extensively to biological membranes and enhances lipid peroxidation [9]. Vitamin E (α-tocopherol) as the most important lipophilic antioxidant is a natural component of the membrane lipid bilayer of cells and thus preserving membrane integrity. It inhibits free radical formation and may effectively minimize lipid peroxidation resulting oxidative stress in biological systems because of its oxygen scavenging effect [8,10]. Several studies have been investigated on the effect of chronic administration of DZN and vitamin E for 14 and 21 days on biochemical parameters and/or antioxidant defense status in rat liver and erythrocytes [11], mice liver [12] and rat heart [13]. However, few studies have been performed on acute exposure of DZN and combination of vitamin C and vitamin E [14,15]. To the best of our knowledge, there is no report on the prophylactic and therapeutic roles of vitamin E on DZN-induced oxidative stress in vital tissues after intraperitoneal injection. In the present study, we tried to look at these effects with assessing the activities of free radical scavenging enzymes such as superoxide dismutase (SOD), catalase (CAT), glutathione S-transferase (GST), as well as malondialdehyde (MDA) level as an important index of lipid peroxidation and glutathione (GSH) concentration in liver, erythrocytes, kidney, brain, heart and spleen and biochemical parameters measurement in plasma of male Wistar rats.
Materials and Methods
Chemicals
DZN, purity 99% was obtained from Supelco Company (USA). All chemicals were of the highest analytical grade from Sigma and Merck. DZN and vitamin E dissolved in corn oil at a stock concentration of 600 mg/mL, immediately before use.
Animals
Male Wistar rats weighing 170–230 g were purchased from Baqiyatallah University of Medical Sciences (Tehran, Iran) and kept for 1 week before the beginning of the experiment for acclimatization. Animals were housed three to a cage at room temperature (25±2 °C) with a relative humidity of 50–60% and on a 12 h light–darkness cycle. The animals had free access to commercial pellet diet and water ad libitum. The ethics committee of the Baqiyatallah University of Medical Sciences approved the experimental protocol, and all efforts were made to minimize the animal suffering.
Experimental design
The animals were randomly divided into five groups having six rats in each group. Control group was treated with corn oil as DZN solvent. DZN group was treated with 100 mg/kg of DZN [7]. E group was treated with 150 mg/kg of vitamin E [8]. Rats in E-DZN group were pre-treated with 150 mg/kg of vitamin E and then dosed with DZN (100 mg/kg), 30 min later. Rats in DZN-E group were pre-treated with 100 mg/kg of DZN and then dosed with vitamin E (150 mg/kg), 30 min later. All injections were carried out intraperitoneally. All rats were weighed at the beginning and at the end of the study.
Plasma and tissues preparation
Rats in each group were sacrificed 24 hours [7,16] after the last administration following overnight fast. Blood samples were collected by cardiac puncture into heparinized test tubes, and immediately centrifuged at 3,000×g for 15 min at 4 °C. Plasma was removed and erythrocytes were washed three times with five volumes of phosphate-buffered saline (PBS), centrifuged as above, and after removal of the supernatant fluid and the white buffy layer, were divided into equal portions and frozen at −70 °C until use. Also, liver, kidney, brain, heart, and spleen were quickly removed, washed in ice-cold PBS. Washed tissues were immediately immersed in liquid nitrogen and stored at −70 °C until biochemical analysis.
On the day of use, erythrocytes were hemolyzed in 10 volumes of ice-cold distilled water. After centrifugation, the supernatant was used for biochemical analysis. In addition, frozen tissue samples were quickly weighed and homogenized 1:10 in ice-cold PBS in a Heidolph type homogenizer (Germany). The homogenates were then centrifuged at 16,000×g for 15 min at 4 °C. The supernatants were separated and used for enzyme activities assays and determination of GSH, MDA and protein levels.
Plasma biochemical parameters assays
Activities of aspartate transaminase (AST), alanine transaminase (ALT), alkaline phosphatase (ALP), lactate dehydrogenase (LDH) and creatin phosphate kinase (CK), levels of urea, creatinine and uric acid were determined in plasma using Parsazmun Company kits (Tehran-Iran).
ChE activity assay
ChE activity in cell lysates was determined at 412 nm by a slight modification of Ellman method [17]. The reaction mixture containing 0.05 mL homogenate and 0.423 mM 5, 5′-dithiobis 2-nitrobenzoic acid (DTNB) in 0.1 M sodium phosphate buffer, pH 7.5 was incubated at 37 °C for 5 min. Reaction was initiated by addition of 20 mM acetylthiocholine iodide as the substrate for the erythrocytes, brain and spleen cells and butyrylthiocholine iodide for the liver, kidney and heart. The rate of increase of absorbance was measured on a Genesys 10 UV spectrophotometer at 412 nm during 5 min. The enzyme activity is expressed as U/ mg protein.
Biomarkers of oxidative damage
The activity of SOD was determined based on the ability of SOD to inhibit the reduction of nitroblue tetrazolium by superoxide. CAT activity was measured at 240 nm by calculating the rate of degradation of hydrogen peroxide (H2O2) as the substrate of the enzyme. GST activity was assayed by monitoring the formation of the thioether product of the reaction between GSH and 1-chloro-2, 4-dinitrobenzene at 340 nm. GSH level was assayed by monitoring the absorbance of DTNB at 412 nm. MDA level as an indicator of lipid peroxidation was determined at 532 nm using 2-thiobarbituric acid [7,18].
Protein and hemoglobin levels
The total protein concentration in tissues was measured by Bradford’s method [19]. Appropriate volume of sample reached the volume of 1 mL to which 3 mL of Bradford solution was added and was incubated for 10 minutes. Then, absorbance was read at 595 nm. Protein concentration was calculated using 1mg/mL bovine serum albumin solution as standard.
Hemoglobin (Hb) concentration in blood was measured by Van Kampen method [20]. 100 μL of whole blood samples were mixed with 5 mL of Drabkin’s solution and the hemoglobin concentration was measured with a hemoglobinometer (ERMA INC, Japan). Protein and hemoglobin concentrations were used to normalize enzyme activities and levels of GSH and MDA.
Statistical analysis
Statistical analysis of the data was conducted using SPSS statistical software version 22 (IBM Corporation, USA). Significance was determined by analysis of variance (ANOVA) followed by post hoc analysis using Tukey multiple comparison tests applied across treatment groups for each tissue. Analysis of the correlation between oxidative stress biomarkers and ChE activity was performed using Pearson correlation analysis. Results were expressed as mean±SD. Significance level was based on p<0.05.
Results and Discussion
Effects of treatments on plasma biochemical parameters
The increase in the activities of ALT, AST, LDH, ALP and CK enzymes and urea, uric acid and creatinine levels are used as important diagnostic tool for cellular damage and cytotoxicity of toxic agents [21–23]. In this study, DZN significantly increased plasma AST, ALT, ALP, CK and LDH activities and urea, uric acid and creatinine levels compared with those of the control group (p<0.05) (Figure 1 and 2). The elevated these enzymes activities in DZN exposed rat may be due to leakage from the liver and heart tissues as a result of DZN-induced lipid peroxidation of membrane [24]. In addition, significant increases of plasma creatinine, urea and uric acid levels may due to the impairment of the glomerular function and tubular damage in the kidneys caused by DZN through induction of oxidative stress [2,5]. Similarly, ROS produced from the metabolism of DZN may damage other tissues such as heart, kidney and spleen causing the leakage of enzymes from the tissues into plasma [21]. However, the reduced these parameters levels in groups pre- and post-treated with vitamin E showed that the antioxidant vitamin E protected the tissues from the lipoperoxidative changes provoked by DZN, probably due to its free radical scavenging ability [2,25]. Damodar et al. [26] have reported that co-administration of vitamin E with DZN to rats resulted in insignificant improvement of the liver enzyme activities. Altuntas and Delibas [27] observed that vitamin E did not show effect on some biochemical indices.
Effects of treatments on ChE and antioxidant enzymes activities
The most important action of DZN compound toxicity is the inhibition of ChE activity and the accumulation of ACh in the peripheral and central nervous system leading to increased activation of nicotinic and muscarinic receptors [28]. In the present study, in vivo administration of DZN inhibited ChE activity in all tissues of rat and administration of vitamin E in the protective and therapeutic groups alleviated the DZN toxicity and maintained the activity of ChE near to the normal value of control in the liver, kidney, brain and spleen (Table 1). This may be due to increase the activity of paraoxonase by vitamin E [29], which is essential in the detoxification of OP and also aids in the reactivation of AChE [30]. Similar results were shown by previous studies [5,31]. A study showed that after 4 weeks of DZN administration, a reduction in plasma ChE activity was observed and vitamin E did not affect the ChE activity [32].
Oxidative stress occurs in the tissues by DZN exposure through increased production of ROS and/or decreased capacity of antioxidant defense. The antioxidant enzymes including SOD and CAT protect cells against the deleterious effects of oxidative stress by converting free radicals to non-radical products. SOD converts superoxide anion radicals into H2O2 and O2. Then, H2O2 is converted into H2O and O2 by CAT [7,33,34]. GST catalyses conjugation of GSH with DZN or ROS and produces more water soluble and excreteable compounds. Therefore, it can decrease the toxicity of DZN and protects tissues from lipid peroxidation and oxidative stress [10]. In this study, DZN increased SOD and GST activities in all tissues and CAT activity in liver, kidney and heart. It also decreased CAT activity in erythrocytes, brain and spleen. There were significant differences in SOD and CAT activities in brain between DZN group and E-DZN group (Table 1). The increased of these antioxidant enzymes activities are probably a response to neutralize the impact of increased ROS generation in tissues [34]. The depletion of CAT activity may be a consequence of irreversible inactivation of enzyme proteins from increased ROS production resulting from DZN metabolism. The increased GST activity was associated with an increase in GSH consumption (Figure 3) indicates the increase of body’s defense against DZN, and rapid excretion of it [35]. However, pre and post-administration of vitamin E had the ability to recover these parameters due toits ability to scavenge the accumulated free radicals leading to limiting the effects of ROS on the tissues, and thus it protects the cell from injury [11,36,37]. Vitamin E is a nonenzymatic antioxidant in the lipid phase of cells, acts to neutralize the toxic effects of ROS, which are potentially damaging by-products of the body’s metabolism [8]. Previous studies performed in animals support our study findings. Ibrahim [37] reported that SOD and CAT activities were significantly increased in kidney and liver tissues of fish after 14 and 28 days exposure to DZN, while these enzymes were significantly decreased in DZN-exposed fishes fed with diets supplemented with vitamin E. Akturk et al. [14] showed that DZN caused significant increases in the activities of SOD and CAT in the kidney of rat and the treatment with a combination of vitamins E and C 30 min after the administration of DZN was somewhat effective in restoring the activities of these enzymes. Messarah et al. [11] indicated that CAT activity was significantly decreased in rat liver and erythrocytes after DZN orally administration for 21 days and the supplementation of vitamin E significantly modulated the activity of this enzyme. El-Shenawy et al. [12] showed a significant decrease in SOD and CAT activities in liver by DZN-exposed mice for 14 days and these effects were largely prevented by a vitamin E supplementation. However, Yilmaz et al. [8] found that only the activity of SOD was directly mediated by DZN exposure and co-treatment with vitamins E and C.
Effects of treatments on GSH level
GSH plays a crucial role in intracellular protection against ROS. It is involved in maintenance of other antioxidants, such as ascorbate and α-tocopherol. GSH is also required for the antioxidant enzymes activities such as glutathione peroxidase and GST [7,38]. In the current study, GSH level was significantly decreased in all tissues of DZN-treated rat comparing to the control (Figure 3). The decreased tissues GSH content indicates impair the cells defense against the toxic actions of ROS, which may lead to oxidative stress and cytotoxicity [2]. However, administration of vitamin E with DZN in the prophylactic and therapeutic groups were returned GSH level near to the control levels, which can be directly correlated to the scavenging of oxidative free radicals. Our previous studies had shown that simultaneous treatment of vitamin E to the DZN-exposed rats was somewhat effective in restoring the activity of GST and level of GSH in different tissues [10,39–43].
Effects of treatments on MDA level
MDA is a stable metabolite of the free radical-mediated lipid peroxidation cascade and it is known as a marker of oxidative stress [22,44]. Our results showed a significant increase in the level of MDA in all tissues except spleen (Figure 4), which might have resulted from oxidative damage induced by DZN, might be due to its lipophilicity, whereby it could penetrate easily to the cell membrane and caused membrane lipid peroxidation [9]. However, pre and post-administration of vitamin E resulted in a significant reduction in lipid peroxidation in DZN treated rats due to the potency of vitamin E to scavenge the free radicals, which are generated during the oxidation of unsaturated lipids and lead to the propagation of lipid peroxidation [12]. Vitamin E allow free radicals to attract a hydrogen atom from the antioxidant molecule rather than from polyunsaturated fatty acids, thus breaking the chain of free radical reactions, the resulting antioxidant radical being a relatively non-reactive species [45]. In several studies, chronic administration of DZN with vitamin E only or in combination with vitamin C significantly decreased MDA level in various tissues [8,11,12,37].
Analysis of correlation between brain oxidative stress biomarkers and ChE activity
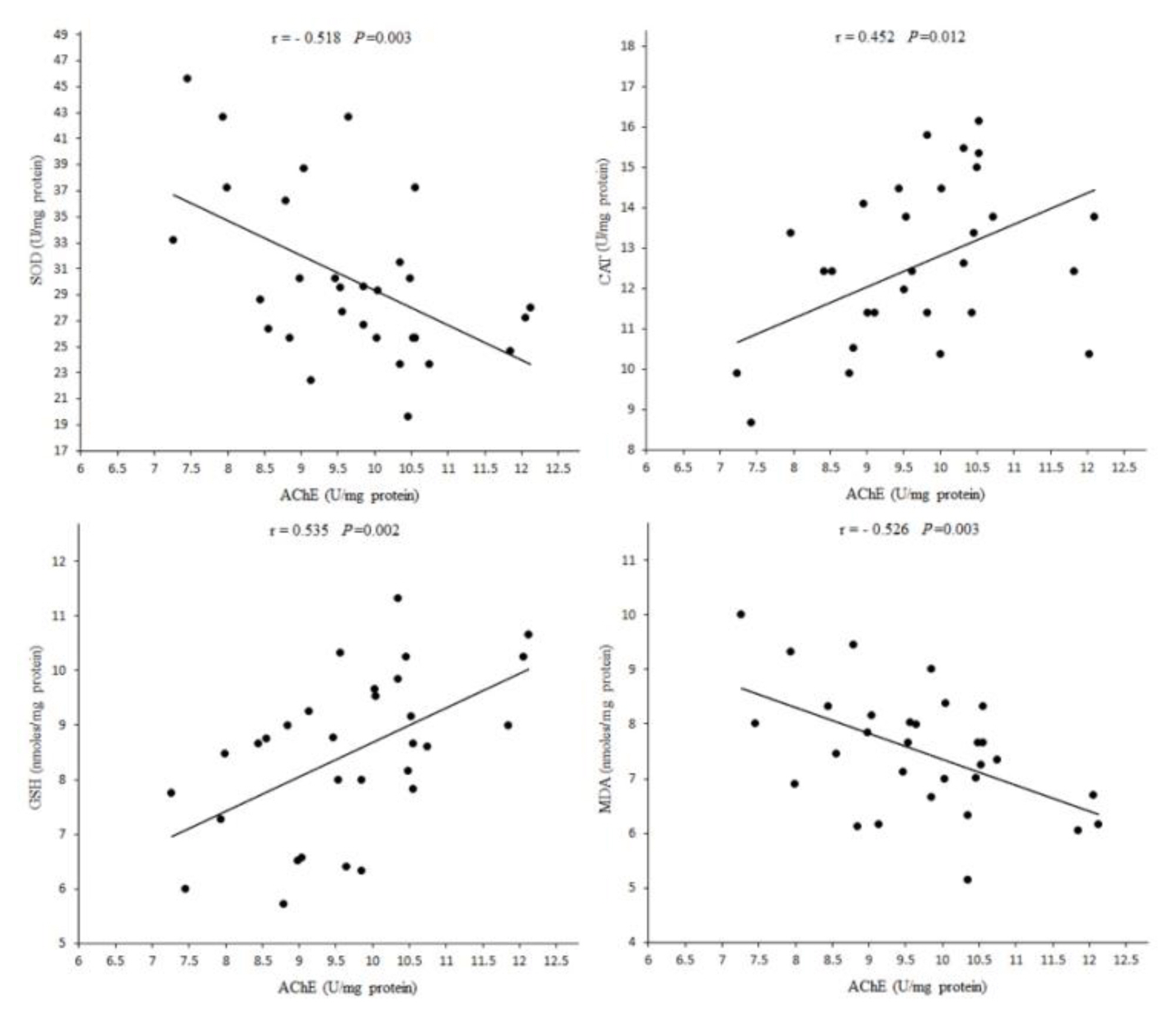
Due to changes in oxidative stress biomarkers in tissues, in the present study, the toxicity of DZN in brain tissue is higher than other tissues. High ratio of membrane surface area of brain to cytoplasmic volume, high content of polyunsaturated fatty acids and the presence of Fe and oxygen, contribute to the accumulation of lipid peroxidation products during oxidative stress induced by DZN products [2,21]. AChE activity in brain was positively correlated with CAT activity and GSH level and negatively correlated with SOD activity and MDA level (Figure 5), suggesting that increased oxidative stress and lipid peroxidation by DZN may be a result of cholinergic hyperactivity which may be an initiator of accumulation of free radicals and cell injury [2]. By comparing the effects of vitamin E on oxidative stress biomarkers in various tissues, it seems that the effects of the restoring in the brain are higher than other tissues.
Conclusions
The findings of the present study suggest that oxidative stress may be involved in DZN toxicity. However, both prophylactic and therapeutic treatments of rats with vitamin E is able to ameliorate the acute toxic effect of DZN on some biochemical and oxidative stress indices through inhibiting the ROS and lipid peroxidation generation, preventing the decline of antioxidant defense system and scavenging free radicals (Figure 6).