Introduction
In agriculture, pesticides are widely used to control pests that reduce yield in order to boost productivity [1]. Cypermethrin and dimethoate are common pesticides used to control pests that affect crops (rice, cotton, soyabeans and vegetables) and animals (sheep, cattle and poultry) [2,3]. Cypermethrin (C22H19Cl2NO3, CAS No. 52315-07-8), is a pyrethroid pesticide frequently used to control acarids and insects [4]. The mode of action is primarily by inactivation of the voltage-sensitive sodium channel of the pest’s neural membrane [5]. This allows more sodium ions to cross and depolarize the membrane beyond the normal level. The consequences of this action are T-syndrome (aggressive behavior, hyper-excitation, ataxia and whole-body tremor) and CS-syndrome (hypersensitivity, excessive salivation, clonic seizure and choreoathetosis), which results in paralysis and death [5]. Dimethoate (C5H12NO3PS2, CAS NO 60-51-5) is an organophosphate pesticide widely used in agriculture to control broad range of insects. The mode of action is phosphorylation of serine residues of the active site of acetylcholinesterase (AChE), which results to inhibition of AChE [6,7] and subsequent overstimulation of effector organs by acetylcholine. Ultimately, nervous system failure, impairment of the respiratory myocardial and neuromuscular transmissions and death occur [7–9].
A study on the efficacy of cypermethrin and dimethoate showed that cypermethrin acts immediately on application and remains effective for a long time, while dimethoate shows a lag period before attaining maximum potency for a short period [10]. Pesticides containing these chemicals can achieve faster and long lasting actions at the same time. Cypermethrin and dimethoate therefore potentiate the efficacy of either chemical or other pesticides. Understandably, some commercial preparations contain these two pesticides as the additive or synergistic effect of the chemicals enhances potency for effective and efficient elimination of pests. These pesticides are frequently applied in rice paddies along river floodplains in Nigeria. Incidentally, these floodplains also serve as breeding sites for some economically important fishes such as Clarias gariepinus [11]. Over 99% of applied pesticides remain in the ecosystem [12] and through run-offs and atmospheric dropout, these can reach surface waters close to agricultural lands where fish breed [13,14]. Persistence of the pesticides in these breeding sites may predispose the juvenile fish populations and other aquatic organisms to toxicity effects [3,15–17]. Reports have shown that pesticides affect fish at ecosystem, population, organismal and sub-organismal (system and organ functions) levels [18].
Uptake of pesticides from the environment by fish species is common [16] via direct uptake from the water and through the food web [13]. Most pesticides are fat soluble and easily bioaccumulate in fish [19]. Mahboob et al. [13] reported cypermethrin concentrations of 0.35–3.47μg/kg in Catla catla from a pesticide polluted river. On uptake, the pesticides may be bio transformed, metabolized and then eliminated or bioaccumulated [5]. Bioaccumulated pesticides could damage some vital organs in the fish [15,20] and cause severe ecological damages [6]. Furthermore, metabolism of most pesticides and xenobiotics in fish results in the production of reactive oxygen species (ROS) [1,4,21]. This could lead to oxidative damage of the liver, kidney, gill, brain, blood, heart and muscular cells [4,7,8,22,23]. Such damages could impair the functions of the affected organs, alter fish behavior and cause death of the fish [8]. Available information on cypermethrin and dimethoate shows that individually, these chemicals alter fish behavior, affect blood parameters; total erythrocyte and leucocyte counts, hemoglobin and mean corpuscular volume [2,7,8,24] and damage cells of essential organs (gills, liver, kidney and brain) [2,20].
Based on the mode of actions, the toxicity of combinations of cypermethrin and dimethoate should be higher than either chemical. However, studies on the combined effect of cypermethrin and dimethoate are rare though these chemicals are combined in some commercially available pesticides and are also commonly found together in aquatic ecosystems [25]. Sangchan et al. [25] observed concentrations of 0.2–0.4 μg/L and 0.01–0.5 μg/L of dimethoate and cypermethrin, respectively in a river close to an agricultural area. Higher concentrations (0.54–0.71 μg/L) of cypermethrin was observed in a stream water in Argentina [26].
Laboratory ecotoxicological studies are designed to provide essential information on the toxicity of xenobiotics to target and non-target organisms in nature [10]. Consequently, for accurate prediction, experimental conditions must be near similar to natural exposure. Since oftentimes rice farmers apply these pesticides once during a planting season, we designed a static non-renewal system to evaluate how juvenile catfish may respond to the toxicants. Therefore, this study aims at evaluating the effects of different combinations of cypermethrin and dimethoate on the behavior, hematology and histopathology of juvenile C. gariepinus. The study will provide additional information that will assist in understanding the toxicity of combinations of pesticides on non-target species. Such information is essential in modeling the effects of concomitantly occurring pesticides on aquatic organisms and deciding regulatory policies.
Materials and Methods
Experimental fish and chemicals
Juvenile C. gariepinus (mean weight of 9.3±0.27 g and mean total length of 11.8±0.27 cm) procured from a fish farm were acclimated for two weeks in the laboratory before commencement of the experiment. During acclimation, the fish were fed daily with commercial catfish food at 2% of their body weight. The pesticide used for the experiment is a commercial formulation of cypermethrin (30 g/L) and dimethoate (250 g/L) with the trade name “Superplus 280 EC”.
Experimental design
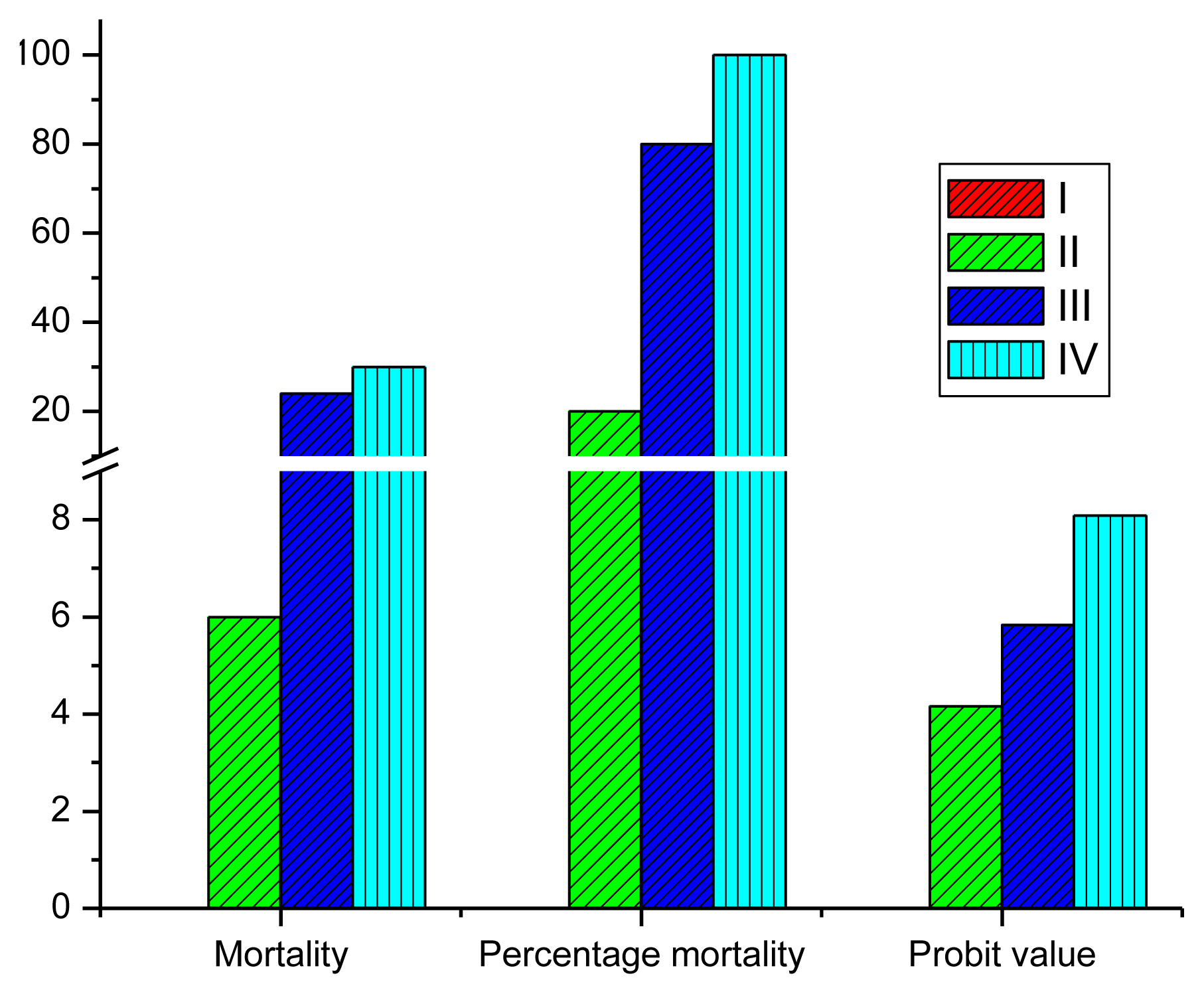
After the acclamation period, series of tests to determine the range of toxicant concentrations that produced a targeted range of effects were conducted. Thereafter, a static lethal toxicity assay was conducted to determine the 96-hour LC50. The choice of static non-renewal experimental design was based on the knowledge that the half-life of most pyrethroid pesticides is longer than 48 hours [27] and the desire to accommodates time-dependent degradation of the pesticides. Four different concentrations of pesticides, 0.75 μg/L cypermethrin and 6.25 μg/L dimethoate (Group II); 0.9 μg/L cypermthrin and 7.5 μg/L (Group III), 1.13 μg/L of cypermethrin and 9.3 μg/L of dimethoate (Group IV) and control, no pesticide (Group I) were used for the lethal assay. The experiment was conducted in triplicate glass tanks (60×30×30 cm) containing 10 fish each in 20 L of water for each group. At the end of the 96-hour experiment, the LC50 was computed and three different concentrations were used to test the sublethal effect of the toxicant. The 96-hour LC50 value derived from probit method was 0.60 μg/L cypermethrin and 5.56 μg/L dimethoate.
Sublethal test
In order to test the sublethal toxic effect of cypermethrin and dimethoate, 120 juvenile catfish were divided into four groups. These are group A (control, without pesticide), group B (0.30 μg/L of cypermethrin and 2.50 μg/L of dimethoate), group C (0.38 μg/L of cypermethrin and 3.13 μg/L of dimethoate) and group D (0.50 μg/L of cypermethrin and 4.10 μg/L of dimethoate). All experiments were performed in triplicate and each tank contained 10 fish in 20 L water. The mean water quality parameters were maintained at total dissolved solids (361.0±2.0 mg/L), conductivity (618.0±4 μS/cm), temperature (27.0±0.1 °C), pH (7.3±0.1) and dissolved oxygen (6.0±0.5 mg/L). Throughout the duration of the experiment, the fish were visually monitored for behavioral changes such as erratic swimming, standing erect, somersaulting, air gulping, sudden jerk movement, rapid opercula movement and release of bubbles. These were virtually assessed daily by subjective comparison between test groups and control.
Hematological analysis
At the end of the 96-hour experiment, blood from five different fish from each tank was collected from the caudal fin using heparinized syringes and transferred to EDTA tubes. Hematological analysis of red blood cell (RBC), packed cell volume (PVC), hemoglobin (HB), mean corpuscular hemoglobin concentration (MCHC), mean corpuscular hemoglobin (MCH) and mean cell volume (MCV) was performed using hematology analyzer.
Histopathological analysis
On completion of the experiment, the fish were euthanized and the liver, kidney, brain, gill and fin were collected for histological analysis. Firstly, they were fixed in 10% normal saline and then dehydrated using 70%, 90% and absolute alcohol for ten minutes each. Secondly, they were dried by immersion in three changes of xylene for ten (10) minutes each. Thirdly, they were impregnated in paraffin wax in a hot oven at a temperature of 60 °C. Thereafter, blocks were made, sectioned at 5 μm thickness using a rotary microtome, rehydrated in distilled water and stained with Hematoxylin-Eosin (H-E). The rehydrated sections were examined and micrographed under light microscope [28].
Statistical analysis
The median lethal concentration that caused 50% mortality of exposed animals (LC50) was calculated using probit analysis based on a computer programme. The statistical difference between test groups was estimated with analysis of variance (ANOVA) using Statistical Programme for Social Sciences (SPSS) software, version 23.
Results
Acute toxicity
The percentage mortality in the exposed fish varied from 0 in the control group to 100 in Group IV (Figure 1). The 96-hour LC50 value derived from probit method was equivalent to 0.60 μg/L cypermethrin and 5.56 μg/L dimethoate.
Behavioral changes in C. gariepenus during sublethal toxicity test
No adverse behavioral change, discoloration or mortality was observed in the control group. The fish in the different concentrations of the pesticides exhibited erratic swimming, rapid opercula movement, release of bubbles, frequent surfacing (gulping of air), loss of equilibrium (somersaulting) and sudden jerk movement (attempts to jump out of the tanks). The severity of these pathological behavioral changes was concentration and time dependent (Table 1).
Hematological response
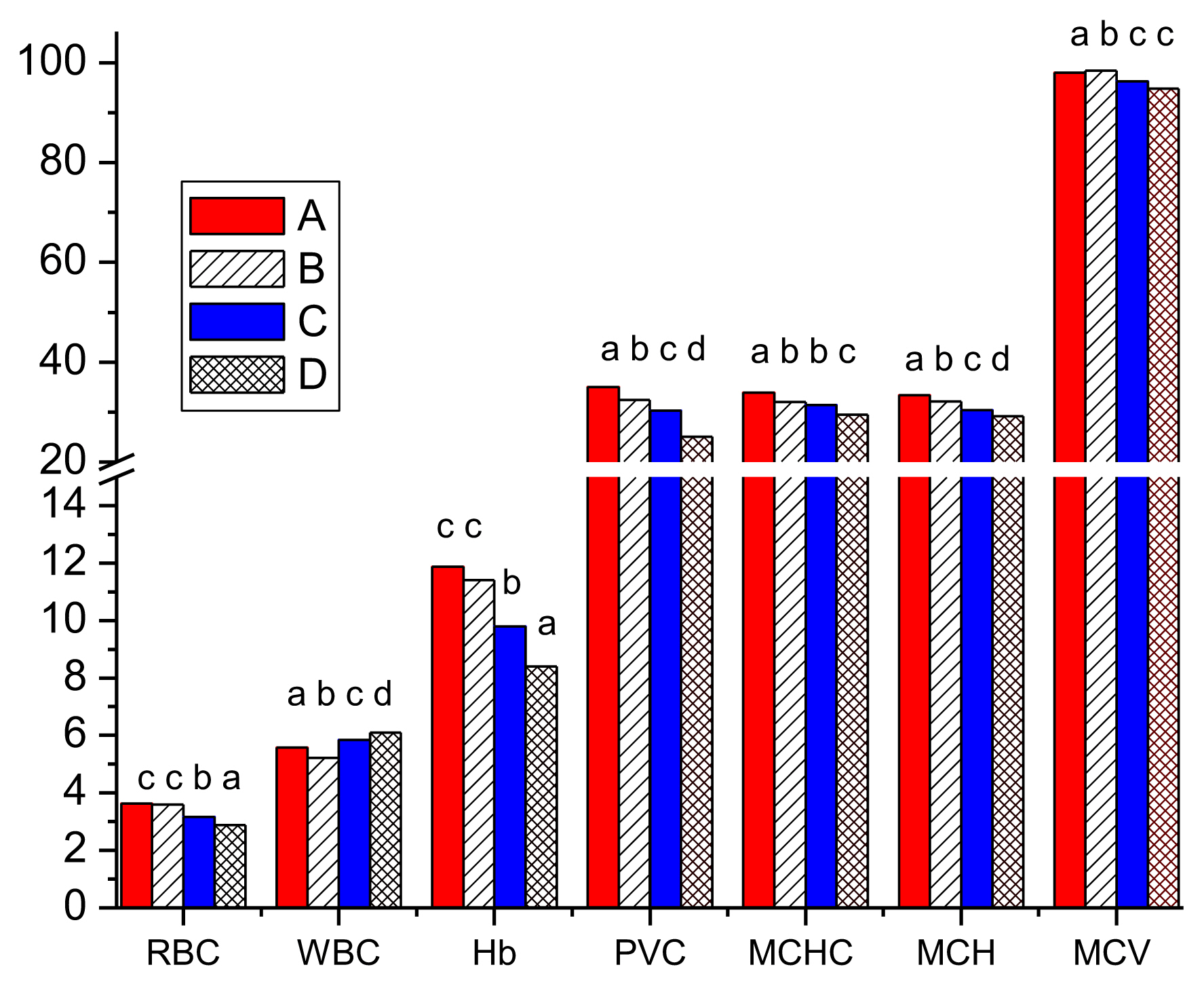
The red blood cells count (RBC) of the control group showed a mean value of 3.63×106 mm−3. The RBC in Groups B, C and D reduced significantly (p<0.05) by 0.83, 12.57 and 20.94%, respectively. A mean white blood cells count (WBC) of 5.58×103 mm3 was observed for the control group, while comparatively significant (p<0.05) increases of 4.84% and 9.32% were observed in test Groups C and D, respectively (Table 2). Contrariwise, mean hemoglobin (Hb) values reduced significantly (p<0.05) by 4.12%, 17.43% and 29.23% in Groups B, C and D, respectively compared to the control. Consistent with Hb, PCV decreased significantly (p<0.05) in a concentration dependent manner, with the lowest mean value (25.00±0.58%) recorded for the highest concentration (Group D). Mean corpuscular hemoglobin concentration (MCHC), mean corpuscular hemoglobin (MCH) and mean cell volume (MCV) followed the same pattern as Hb and PCV with the lowest values in the highest pesticide concentration (Figure 2).
Histopathological response
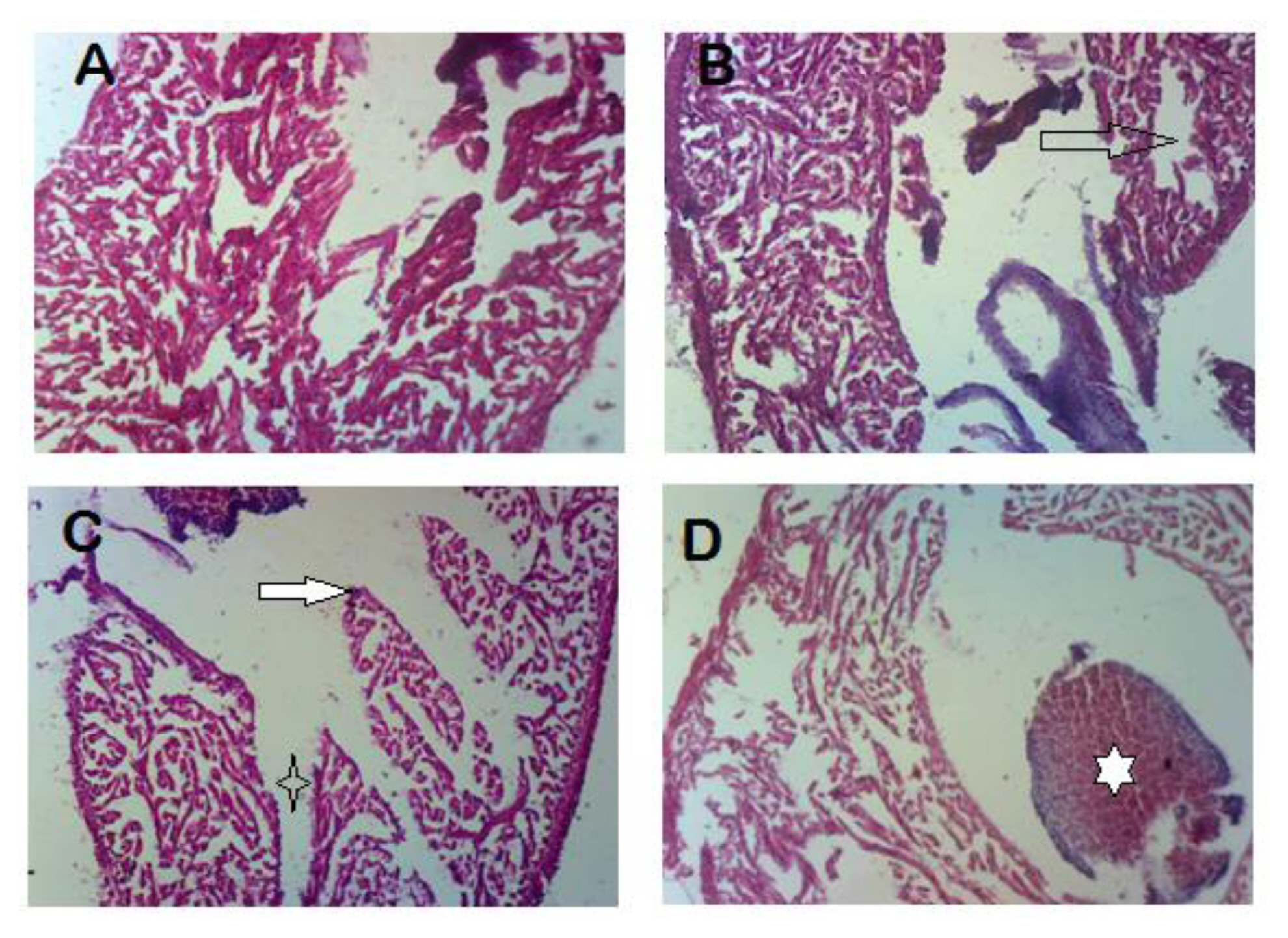
Histopathological examination showed normal cardiac muscles and cardiac chambers in the heart of fish in the control group. Group B showed a disorganization of intra-cardiac chamber structure, while group C showed saw-toothed intra-cardiac chamber muscles with loss of intra-cardiac muscle fiber. Damage to the cardiac muscles was worse in group D with focal loss of cardiac muscle and intra-cardiac hemorrhage (Figure 3).
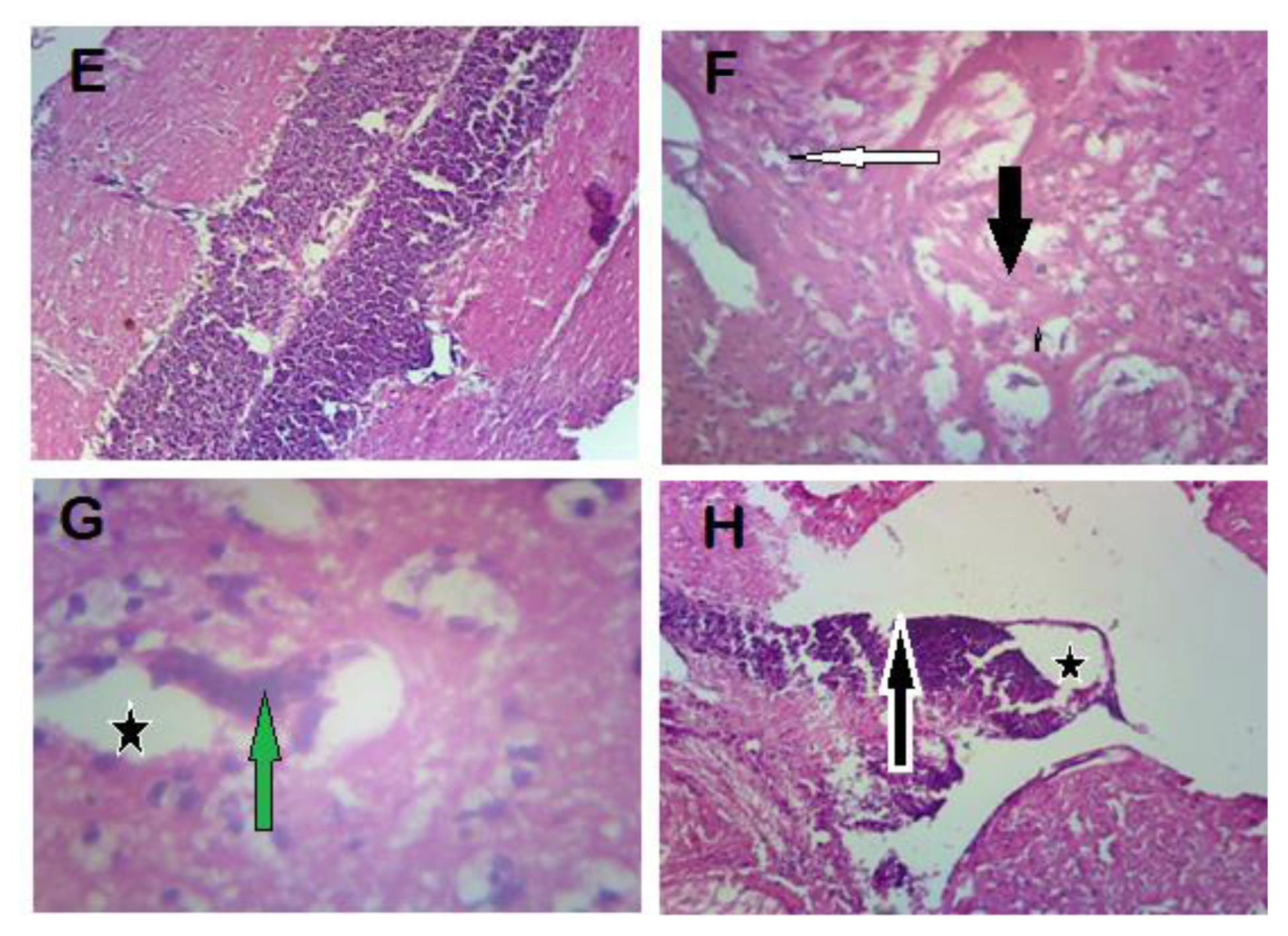
There was no evidence of damage in the brain of fish in the control group but spongiosis and infiltration of inflammatory cells (microglial cells) were observed in group B. Group C showed a focal area of liquefactive necrosis with infiltration of inflammatory cells. In group D, there was severe distortion of the brain architecture, focal area of liquefactive necrosis and loss of brain tissues in the area (Figure 4).
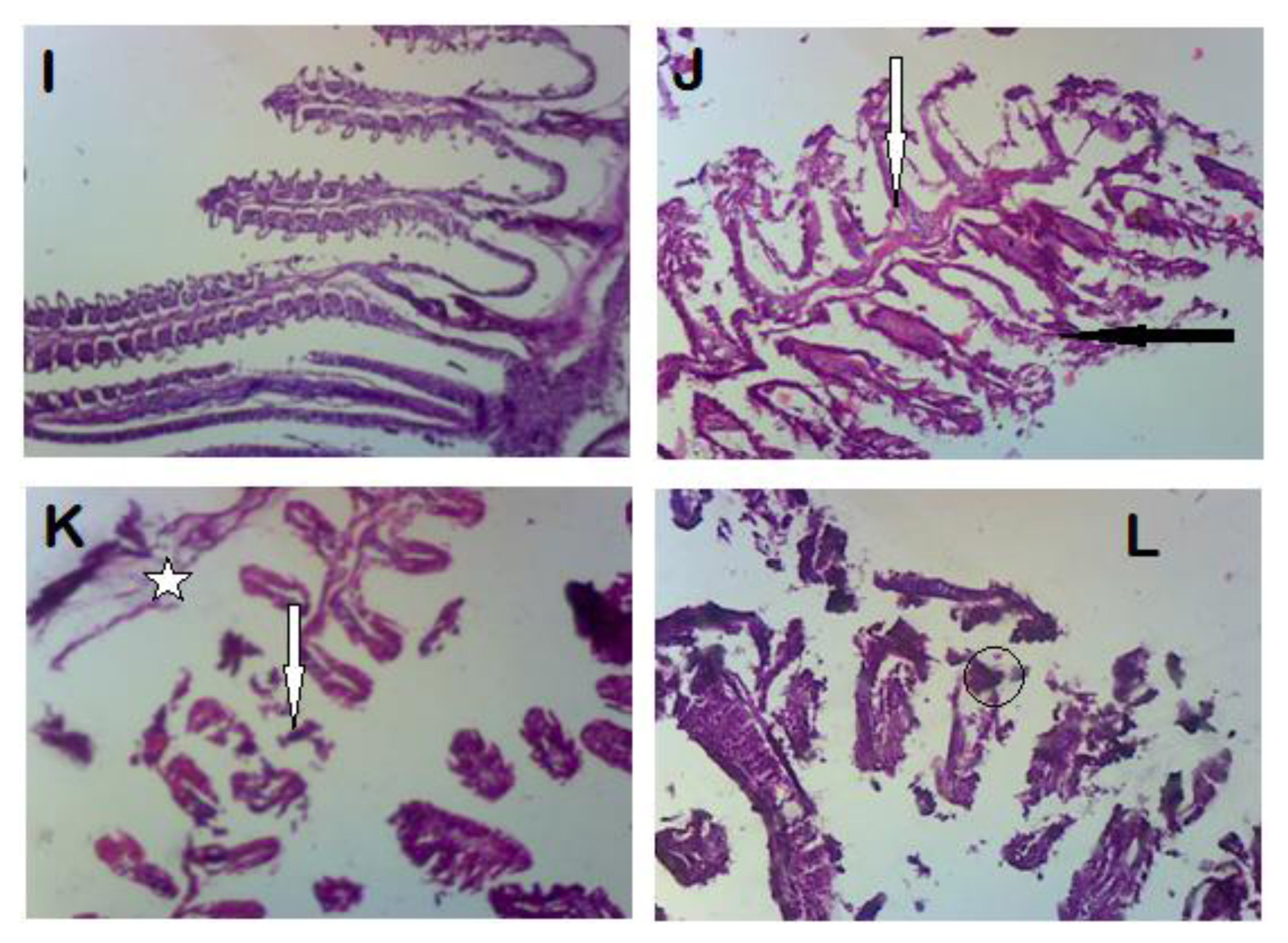
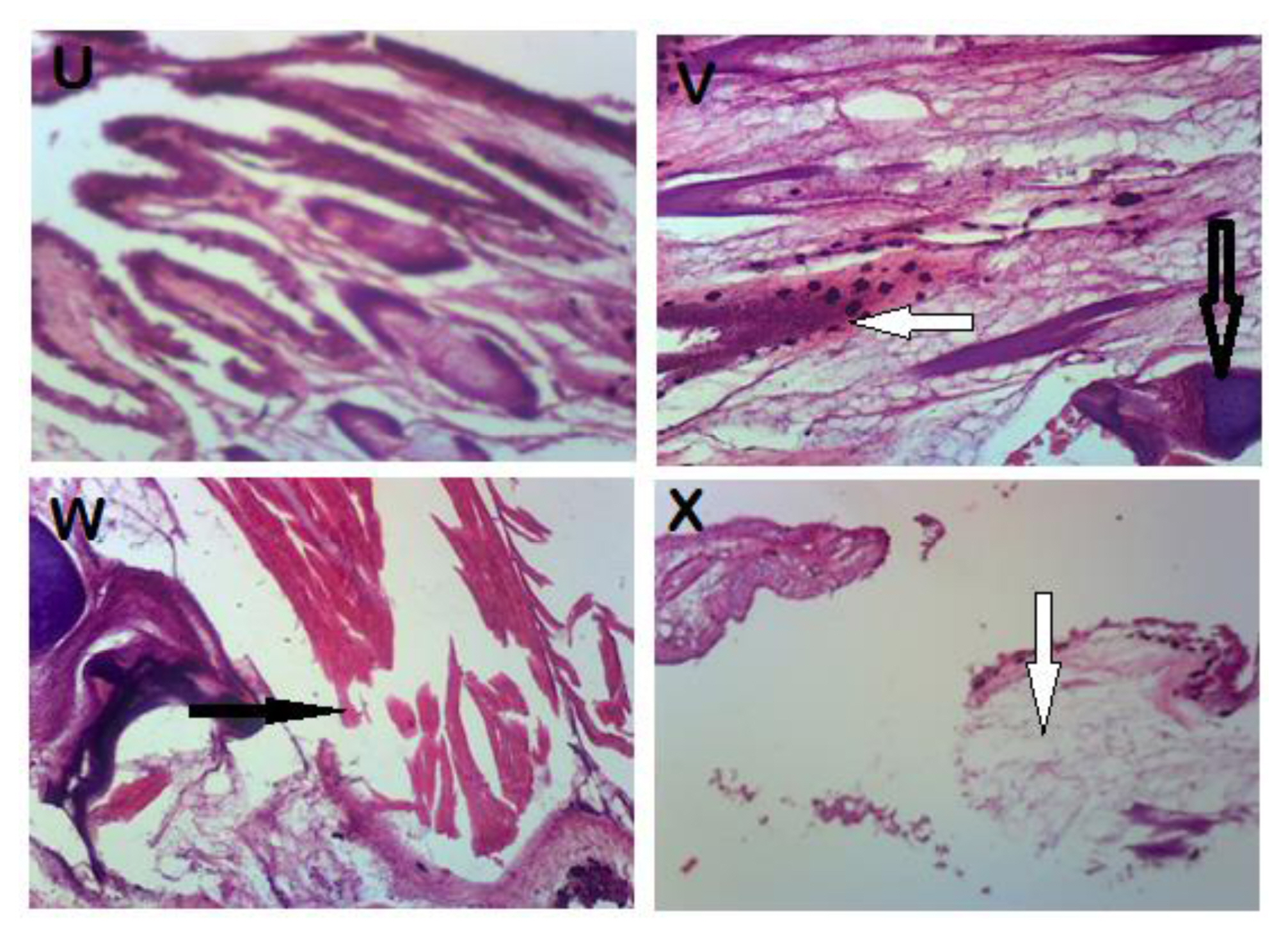
In the gills, histological damages were distortion of gill epithelium, lifting of gill epithelium from the gill stock, disorganization and fragmentation of parts of the gill (Figure 5). Normal hepatocytes were observed in the liver of the control group but aggregation of inflammatory cells, and necrosis and distortion of hepatic architecture were observed in the pesticide exposed groups (Figure 6). The kidney of C. gariepinus exposed to different concentrations of the pesticide showed distortion of cell architecture, fragmentation, loss of renal tissues and intra-renal hemorrhages (bleeding in the renal cells). The severity increased as the concentration of the toxicant increased (Figure 7). Histology of the fin of fish from group B showed fragmentation in the fin and separation from the attached muscles, while the fin from group C shows focal destruction and loss of fin materials or components (Figure 8).
Discussion
The LC50 of the combined pesticide was equivalent to 0.6 μg/L of cypermethrin and 5.56 μg/L of dimethoate. These values are within the lower limits when compared to previous reports on the toxicity of individual chemicals to different fish species. For instance, Velisk et al. [2] reported a 96-h LC50 of 34.1 μg/L for rainbow trout, Saha and Kaviraj [29] reported 0.67 μg/L for H. fossillis, Fai et al. [30] reported 9.13 μg/L and Velmurugan et al. [31] reported 100.4 μg/L for C. gariepinus for cypermethrin only. Amaeze et al. [24] reported LC50 range of 7.60–10.64 μg/L for C. gariepinus exposed to cypermethrin. Sarkar et al. [32] generally put the LC50 of cypermethrin within the range of 0.4 and 2.8 μg/L. Dogan and Can [8] reported an LC50 of 7.35 mg/L for dimethoate for O. mykiss, Qayoom et al. [33] reported LC50 of 1.1 mg/L for Cyprinus carpio, Yildirim et al. [34] reported LC50 of 45.5 mg/L for Oreochromis niloticus, Narra [35] reported an LC50 of 12.45 mg/L for Clarias batrachus, while Amaeze et al. [24] reported 1.70–2.44 mg/L for C. garipinus. Comparing these reports to the present study shows that the combination of cypermethrin and dimethoate increased the toxicity effect and lowered the LC50 of either chemical. Similarly, Fai et al. [30] observed a higher mortality in O. niloticus exposed to a mixture of cypermethrin and dimethoate than in either chemical.
The observed abnormal behaviors were similar to reported toxicity response of other fish species to either cypermethrin or dimethoate [7,1,20] and signal biochemical, physiological and morphological toxicity [8]. The pesticides are known to inhibit neurotransmission and thus affect coordination in intoxicated organisms [1,5,33]. Cypermethrin and dimethoate inactivate the sodium voltage-sensitive channel of the neural membrane and inhibit acetylcholinesterase, respectively resulting in cascades of activities that lead to nervous failure, over stimulation of effector organs, impairment of respiration and neural transmission [5,7]. These toxicity effects are probably responsible for abnormal behaviors such erratic swimming, sudden jerk movement (due to over stimulation of muscles), frequent air gulping (due to impaired respiratory myocardial) and rapid opercula movement. Behavioral changes were analogous to Oncorhynchus mykiss and Heteropneustes fossilis, respectively exposed to dimethoate and cypermethrin [8,29].
Significant decreases in the RBC, HB, PVC, MCHC, MCH and MCV of toxicant-exposed fish compared to the control in concentration dependent manner could be attributed to anemia. Decline in RBC, MCHC, MCH and MCV suggests stress-induced erythropenia probably caused by hemolysis, inhibition of erythropoiesis and, pathological damage to some tissues and hematopoietic organs as reported by Dogan and Can [8], Akinrotimi et al. [36], Majumder and Kaviraj [37] and Özok et al. [1]. These are similar to hematological toxicity of endosulfan and phosphamidon on Barbus conchonius [38], diazinon on C. carpio [2], dimethoate on O. mykiss [8] and C. batrachus [35], glyphosate on C. gariepinus [15] and cypermethrin on O. mykiss [2]. Observed hematological damages were more severe in this study than studies on either cypermethrin or dimethoate alone as reported by Velisek et al. [2], Dogan and Can [8], Bojarski and Witeska [39], Amaeze et al. [24] and Ghayyur et al. [20]. Decrease in RBC, HB, MCHC and MCH reduces the oxygen carrying capacity of the blood and supply to the muscles [35]. These could be responsible for some behavioral changes such as frequent surfacing and attempts to jump from the tanks, air gulping and regular opening of the operculum. These abnormal behaviors are apparently attempts at increasing oxygenation and avoidance of asphyxiation.
Increase in WBC in the exposed fish could be attributed to the fish’s immune response to ameliorate the stress occasioned by the pesticide assault. Narra [35] observed WBC increase in C. butrachus exposed to dimethoate. However, Velisek et al. [2] reported that a higher concentration of 3.4 μg/L of cypermethrin had no effect on the WBC of O. mykiss. Dogan and Can [8] and Ghayyur et al. [20] observed reduction in the WBC of O. mykiss and Cirrhinus mrigala exposed to dimethoate, while Majumder and Kaviraj [37] and Amaeze et al. [24] observed increase in WBC of Oreochromis niloticus and C. gariepinus exposed to cypermethrin. Narra [35] suggested that in the presence of toxicants, leucopoiesis may increase thus elevating WBC. Impliedly, the higher the level of intoxication, the higher the amount of WBC released, which is purely a defense mechanism against the toxicant. It is therefore clear that the toxicity of cypermethrin and dimethoate increased when in combination than singly.
Histopathological damages to the fish exposed to cypermethrin and dimethoate were disorganization of intra cardiac chamber structure, saw-toothed intra-cardiac muscle, loss of muscle fiber and cardiac hemorrhage. Such changes will impair the functions of the heart. Brain damages were exhibited as spongiosis, infiltration of inflammatory cells, focal area of liquefactive necrosis and distortion of brain architecture. Distortion of gill epithelium, lifting of gill epithelium from gill stock, disorganization and fragmentation (desquamation) were observed in the gills of the exposed fish. These damages though commonly associated with pesticide toxicity [2,15,20,31], appeared more severe. Histopathological damages would impair the normal functioning of the gill, including its capacity to extract oxygen from the aquatic environment and several metabolic activities, which could lead to respiratory failure and death. Impaired capacity to take up dissolved oxygen was probably responsible for changes in behavior such as air gulping and rapid opercula movement.
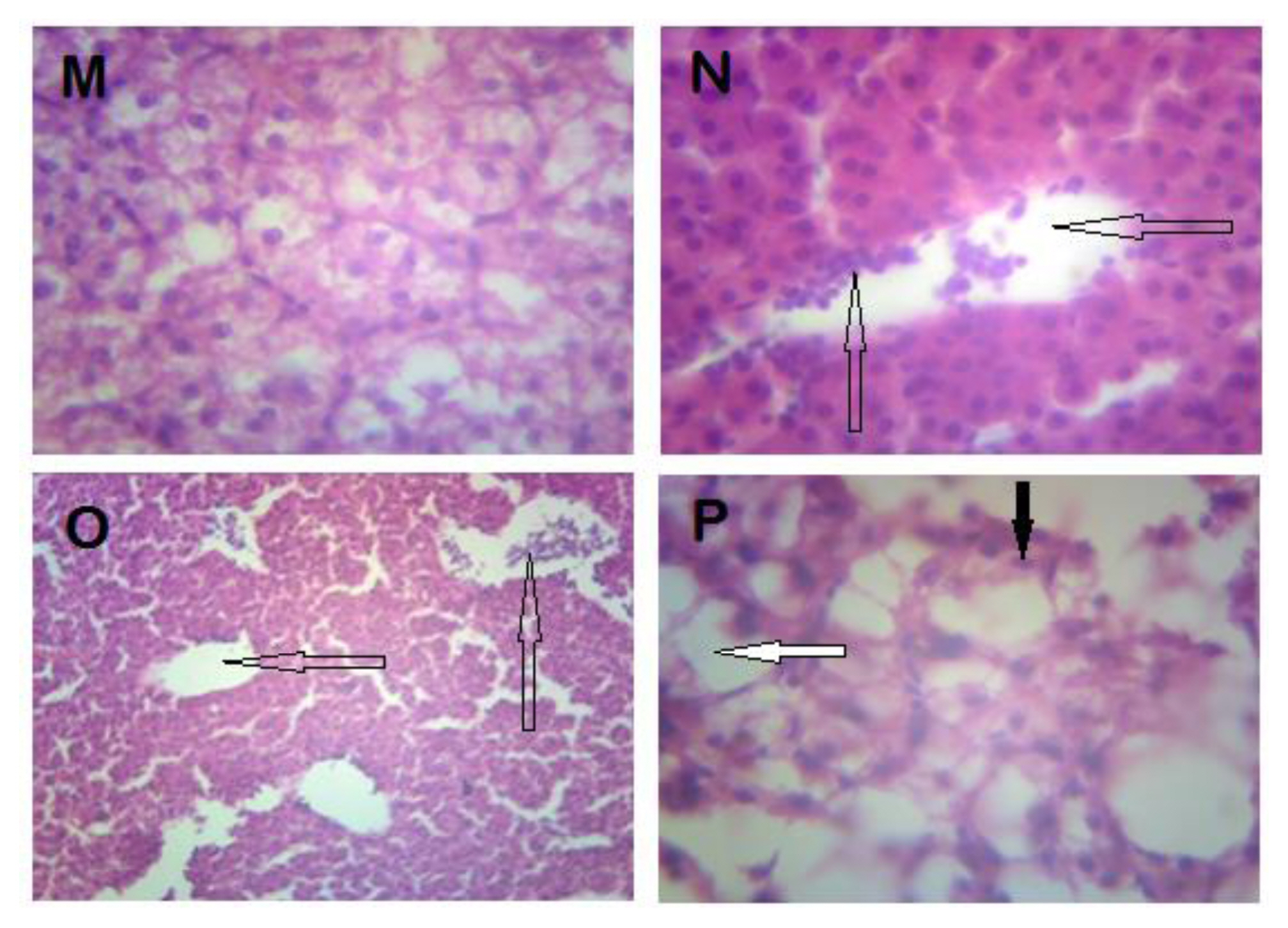
In the liver of exposed fish, distortion of hepatic architecture, focal aggregation of inflammatory cells and necrosis were observed. The liver is the primary organ of detoxification and elimination of xenobiotics, which predisposes it to the toxicity of these toxicants. Observed hepatotoxic effects are consistent with available reports on the toxicity of pesticides to fish [20,31]. Tubular necrosis, distortion of renal tissues and intra-renal hemorrhage were observed in the kidney of exposed fish, which are similar to the findings of [31,34]. In the present study, kidney and gill hemorrhages were probably due to damage to the blood capillaries. This pathological damage is commonly associated with pesticides [28] and explains observed erythropenia. In addition, the mobilization of inflammatory cells in most of the organs provides good explanation for the increase in WBC and such mobilization confirms that it is a response to the toxicity of the pesticide.
Study limitations
Commercially available pesticide, Superplus 280 EC was used in this study and it may contain other chemicals (not listed on the label) that could have affected the results. This study limitation could be circumvented by using pure cypermethrin and dimethoate chemicals, which were not available to us at the time of study. Furthermore, future laboratory studies should include fish from different age classes and species, and time-series field assessment of the toxicants in water and fish in order to provide comprehensive understanding of the environmental and human hazards associated with the pesticides.
Conclusions
The study revealed that juvenile C. garipinus exposed to sublethal concentrations of cypermethrin and dimethoate suffered severe hematological and organ injuries. In combination, the pesticides toxicity increased in time and concentration dependent manner. This is worrisome consequent to the contemporaneity of cypermethrin and dimethoate in the environment and some pesticide formulations. There is urgent need to regulate the use of pesticides containing these chemicals, especially in floodplains that serve as breeding sites for commercially and ecologically important fish species.