Introduction
Agriculture in modern times rely on pesticides in order to control pests that pose a threat to crop productivity [1]. Pesticides may contaminate the soil, air, and water with potential adverse effects on non-target aquatic organisms, plants, mammals and soil microorganisms [2]. They can bioaccumulate in living organisms, biomagnify along the food chain and inadvertently affect human health [3]. Majority of modern-day pesticides are synthetic organic chemicals with a mode of action that interfere with metabolic processes in target organisms [4]. Paraquat (1,1′-dimethyl-4-4′-bipyridinium) and its dichloride salt (1,1′-dimethyl-4-4′-bipyridinium dichloride) are commonly used herbicides in defoliating and drying during the gathering of commercial crops [5]. Similarly, glyphosate (N-phosphonomethyl) glycine (commercially sold as roundup or force up) is a systemic non-selective herbicide which is used extensively, particularly in developing countries for weed control [6]. It acts through the inhibition of amino acid and protein synthesis leading to the death of the weed within days of application. However, the toxicity of glyphosate is potentiated through its formulation with surfactants to facilitate penetration into the weeds. Thus, non-target organisms such as fish may be inadvertently exposed to these glyphosate formulations when applied on land and washed off into nearby surface waters via run-off or spray drift [7] eliciting acute and chronic biological effects to aquatic life [8].
Though, several studies have evaluated the acute toxicity of paraquat and glyphosate singly in various fish species [9] as well as their subacute effects using different biomarkers [8]. Similarly, several studies have evaluated various biomarkers of toxicity of paraquat and glyphosate singly in Nile Tilapia (O. niloticus). Specifically, for paraquat, studies with O. niloticus have reported 96 hLC50 value of 11.84 mg/L [10] with dose-dependent gill pathologies at exposure levels of 12 mg/L and 14.20 mg/L, LC50 value of 20 mg/L as well as liver and gonadal pathologies at 0.5 mg/L at varying temperatures [11] as well as 96 hLC50 value of 17.49 μL/L [12] of paraquat. For glyphosate, studies using O. niloticus have shown varying adverse effects (such as 96 hLC50 value of 16.8 ppm with gill, liver and kidney pathologies [13], gill, liver and kidney pathologies at sublethal concentrations (5 and 15 ppm) [14], alteration in enzyme activities at 1.2 mg/L [15], gill, liver and kidney pathologies [16], nuclear abnormalities [17] at 17.2 mg/L, antioxidant enzymes (SOD, GSH, CAT) activity reduction, among other biomarkers at different concentrations (0.2, 0.8, 4 and 16 mg/L) for 80 days [18], alterations in blood parameters as well as induction of hepatic oxidative stress and DNA damage in the blood at different concentrations (5, 10, 20, 30 and 40 mg/L) Acar et al. [19] of glyphosate.
However, there are scanty studies which evaluate the comparative genotoxic, biochemical and histological effects of the herbicides, paraquat and glyphosate on the Nile Tilapia (O. niloticus) and at the concentrations used in this study. O. niloticus (Linnaeus, 1758) is a deep-bodied fish with cycloid scales. It is a plankton-feeding omnivorous species native to Africa but it is cultured worldwide [20]. Growth retardation and harmful effects at population and community levels may result from micronuclei formation which is marked by alterations in erythrocyte nuclei [21]. Biochemical biomarkers such as catalase (CAT), superoxide dismutase (SOD), glutathione-s-transferases (GST), reduced glutathione (GSH), malondialdehyde (MDA), an index of lipid peroxidation have been used to assess toxic effects in animals exposed to pollutants [22,23]. Histopathological changes can be used as biomarkers of the effects of anthropogenic pollution on organisms and are compatible as indicators of ecosystem health [24].
Consequently, the study aim was to assess the effects of subacute levels of paraquat and glyphosate on genotoxic, biochemical and histological biomarkers in O. niloticus with a view to evaluate potential risk to non-target animals such as fish from diffuse sources of pesticides into aquatic ecosystems.
Materials and Methods
Test compounds
The test compounds utilized in this study were herbicides-paraquat and glyphosate which were purchased from a chemical vendor at Ojota, Lagos, Nigeria. Paraquat (paraforce – active ingredient 200 g W.C. in 1 L) and glyphosate (Force Up-360 g glyphosate/L (in the form of 480 g/L Glyphosate-isopropylamine, salt) [25]. A stock solution of 1 g/L was prepared for each herbicide (paraquat: 5 mL in 1 L of dechlorinated tap water=1 g/L; glyphosate: 2.78 mL in 1 L of dechlorinated tap water=1 g/L) from which working solutions were computed.
Collection and acclimatization of test animal
The Nile Tilapia, O. niloticus (Actinopterygii, Perciformes, Cichlidae) juveniles (length: 15.2–18.5 cm) were purchased from an aquaculture farm in Ayobo, Lagos, Nigeria. They were transported in an open 25 L plastic container to the Environmental Toxicology laboratory at the University of Lagos. The fishes were acclimatized to the laboratory environment in a cylindrical plastic tank (60 L) three-quarter filled with dechlorinated tap water for 7 days [25]. The fishes were fed with Coppens feed (2 mm) twice daily. The dechlorinated tap water was changed every 24 h to prevent buildup of waste metabolites and putrefaction of food materials.
Experimental design for acute toxicity studies of paraquat and glyphosate against O. niloticus
Tests were conducted to obtain the range of concentrations ideal for the definitive tests. Following this, definitive tests were conducted in which five active O. niloticus were randomly introduced into the test tanks containing varying concentrations of paraquat, glyphosate and control (dechlorinated tap water only). Each treatment was duplicated, giving a total of 10 fishes per treatment, including control (without treatment). The concentrations of test compounds were as follows: paraquat-8, 10, 12, 14 and 16 mg/L; glyphosate-0.5, 1.0, 1.5, 2.0 and 2.5 mg/L and control. The bioassay tanks were covered with a net to prevent fish from escaping. Mortality was assessed once in 24 h over the 96 h duration of the experiment [26].
Experimental design for subacute toxicity studies
For the subacute toxicity studies, eight O. niloticus were exposed in triplicates to subacute concentrations (1/10th and 1/100th of 96 h LC50 values) of paraquat and glyphosate as follows: paraquat: 1.10 mg/L and 0.11 mg/L; glyphosate: 0.10 mg/L and 0.01 mg/L and control. The test media were changed every 72 h following a static renewal bioassay procedure [25]. At 14 and 28 d post-exposure, blood was obtained from O. niloticus selected from the test media and control for genotoxicity studies while gills and liver were excised from the same fishes following euthanization [27] for biochemical and histological studies.
Evaluation of genotoxic biomarkers in O. niloticus exposed to paraquat and glyphosate
O. niloticus juveniles (length range: 15.2–18.5 cm) were exposed to subacute concentrations (1/10th and 1/100th of 96 h LC50 value) of the test compounds in triplicates for 28 days. Peripheral blood samples were drawn from the posterior caudal vein [25] with the aid of a 2 mL syringe on day 14 and 28 during the exposure period. For each concentration, at days 14 and 28, three fishes were used with one slide prepared per fish. Thereafter, the blood was smeared on a clean glass slide, fixed using 100% ethanol for 20 mins and dried at room temperature for 24 h [28]. Afterwards, Giemsa (10%) stain was applied to the smear for 10 mins. The glass slides were evaluated under a digital light microscope (Leica® DM500, Wetzlar, Germany) at 1000x for frequencies of micronuclei and nuclear aberrations. At each assessment, 1000 cells per fish were analysed totaling 3000 erythrocytes for each group. For scoring the micronuclei, the criteria of [29] was adopted.
Assessment of biochemical biomarkers in O. niloticus exposed to paraquat and glyphosate
Following the harvest of livers of O. niloticus randomly selected from test and control bioassay tanks, the livers were transported to the University of Lagos Biochemistry laboratory in an ice chest for further analysis. The livers were rinsed in ice cold 1.15% KCL solution, blotted and weighed. Thereafter, the livers were homogenized with 0.1 M phosphate buffer (pH 7.2) in a laboratory mortar. Acid-washed laboratory sand was added and blended in the mortar using a pestle [23]. The homogenate therefrom was centrifuged (2500 rpm) for 15 mins, the supernatant was decanted and stored at −20 °C until analysis. The activities of antioxidant enzymes (superoxide dismutase (SOD), catalase (CAT), glutathione-s-transferases (GST)), reduced glutathione (GSH), and malondialdehyde (MDA) (an index of lipid peroxidation) were determined as described in Sun and Zigma [30], Sinha et al. [31], Habig et al. [32], Sedlak and Lindsay [33] and Buege and Aust [34] respectively.
Assessment of histological biomarkers in O. niloticus exposed to paraquat and glyphosate
Following euthanization at designated post-exposure days, dissected gills and liver of O. niloticus were washed with buffered normal saline. Afterwards, they were fixed for 48 h in Bouin’s fluid, dehydrated through serial (70% to 100%) changes of ethanol, cleared in xylene and embedded in paraffin wax [25]. Thereafter the gills and liver were sectioned at 5 to 6 μm, stained with eosin and haematoxylin then analysed with a digital light microscope (XSZ-801BN model, China) coupled with 12.1 mega pixels camera (Casio, EX-Z450, Japan) [25].
Statistical analysis
The dose-response data for the 96 h toxicity tests was analyzed using probit analysis [35]. One-way analysis of variance (ANOVA) and Least Significant difference (LSD) test was used to analyze statistical differences between the mean of the genotoxicity (micronuclei, binucleated and blebbed cells frequency) and biochemical biomarkers activity in O. niloticus exposed to subacute concentrations of paraquat and glyphosate at days 14 and 28. The results were deemed significant at p<0.05. All analyses were conducted using SPSS 20.0 for windows.
Results
Relative acute toxicity of paraquat and glyphosate against O. niloticus
The median lethal concentration (96 h LC50 value) of paraquat and glyphosate against O. niloticus were 11.20 mg/L and 1.22 mg/L respectively. Glyphosate was estimated to be 9x more toxic than paraquat against O. niloticus.
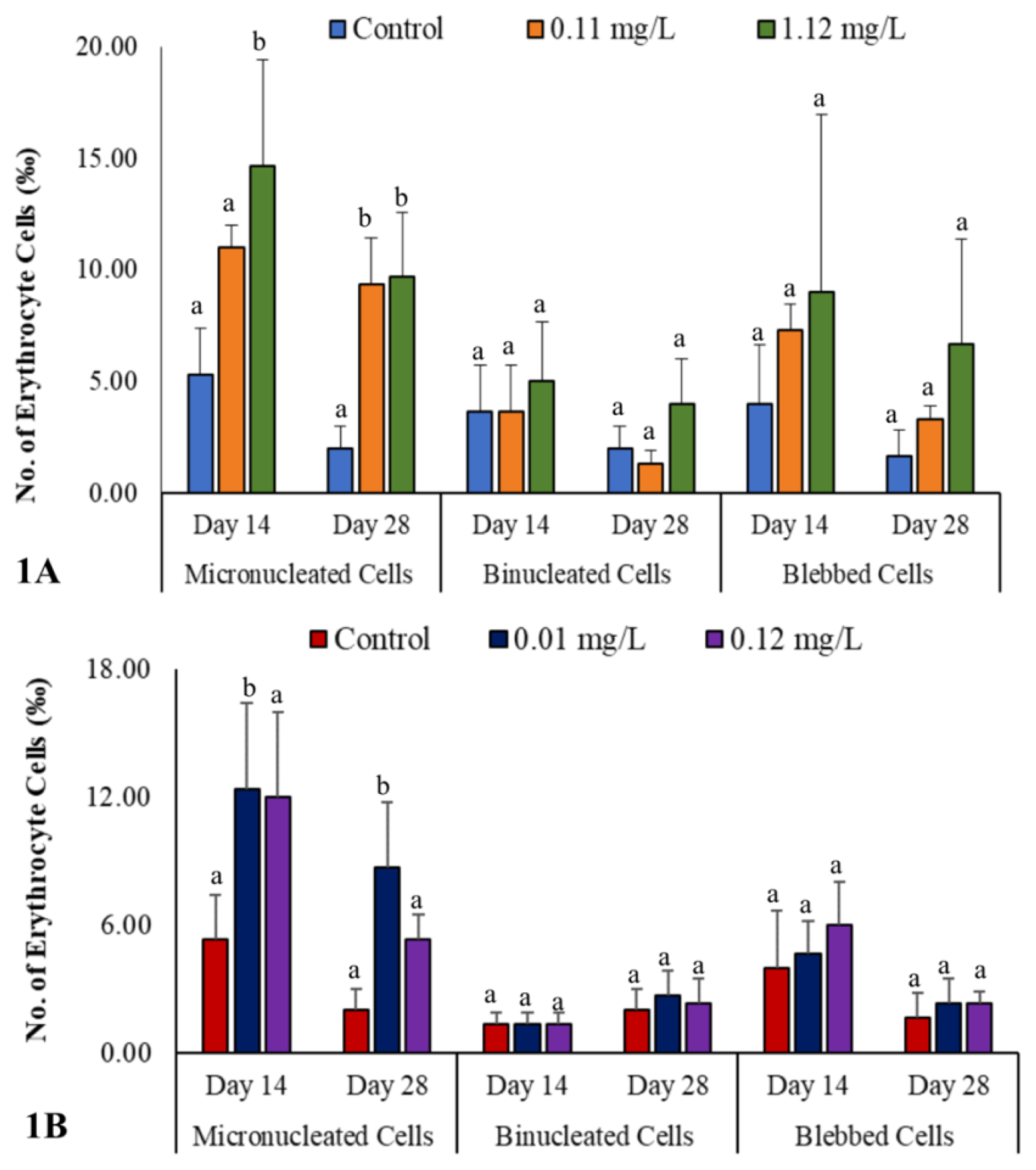
Genotoxic biomarkers in the erythrocytes of O. niloticus exposed to subacute concentrations of paraquat and glyphosate
Binucleated (BN) and blebbed cells were non-significantly higher (p>0.05) at both subacute concentrations of paraquat-exposed O. niloticus compared to control on days 14 and 28. Though, BN cells were non-significantly lower (p>0.05) at day 28 only in the 0.11 mg/L concentration compared to the 1.12 mg/L concentration and control Figure 1A, SI 1. However, micronucleated (MN) cells were significantly higher (p<0.05) at the 1.12 mg/L (higher) concentration only at day 14 and at both subacute concentrations of paraquat-exposed O. niloticus on day 28 compared to control Figure 1A, SI 1.
In glyphosate-exposed O. niloticus, there were no significant differences (p>0.05) between BN and blebbed cells at both subacute concentrations and control on days 14 and 28 Figure 1B, SI 1. However, MN cells were significantly higher (p<0.05) in the erythrocytes of O. niloticus exposed to the lower (0.01 mg/L) concentration when compared to the higher (0.12 mg/L) concentration and control at days 14 and 28 (Figure 1B, SI 1).
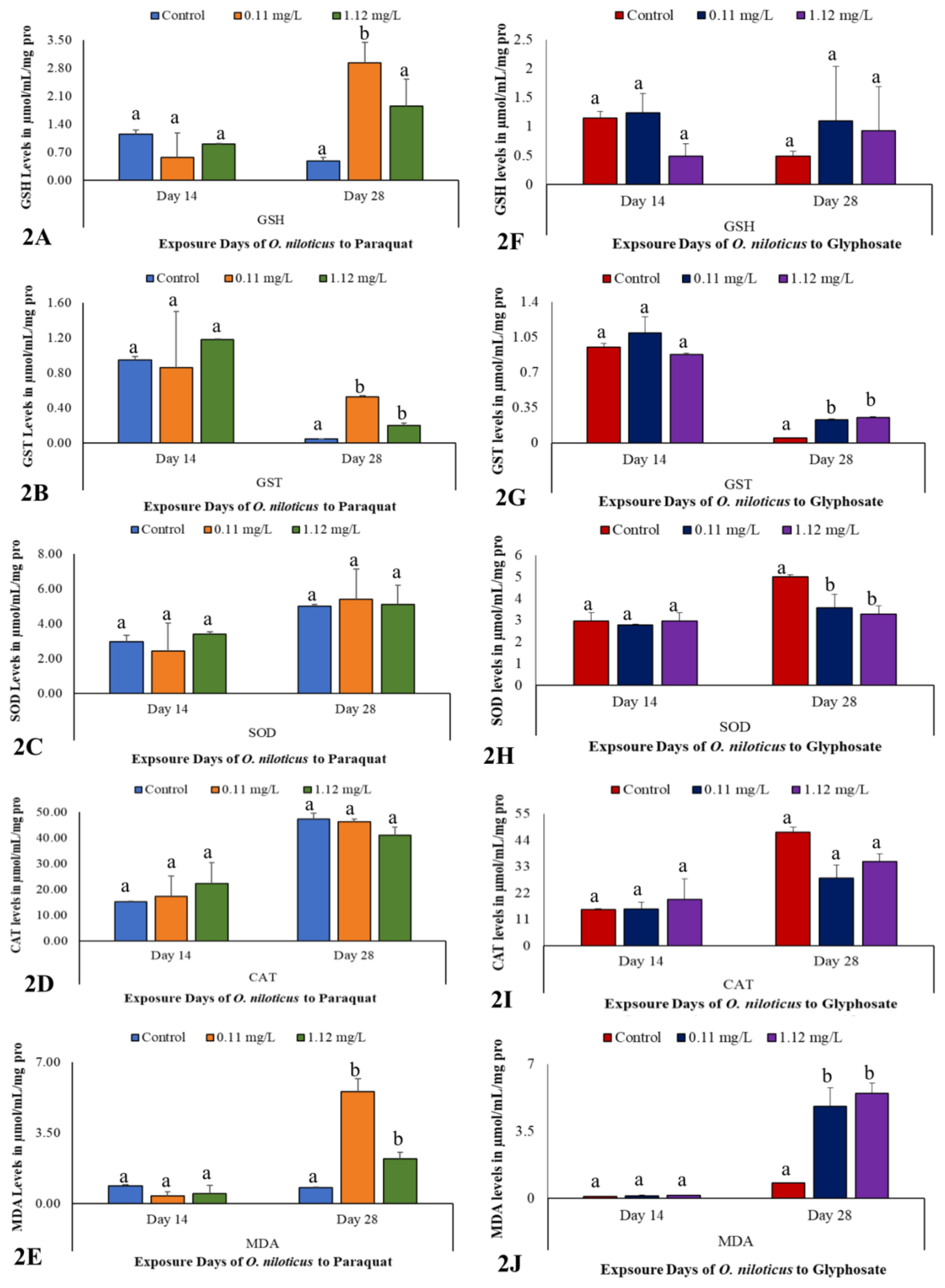
Biochemical biomarkers in the liver of O. niloticus exposed to subacute concentrations of paraquat and glyphosate
At day 14, there were no significant differences (p>0.05) between paraquat-exposed and control O. niloticus as well as glyphosate-exposed and control O. niloticus for all hepatic antioxidant enzymes and MDA activities (Figure 2, SI 2). Also, at day 28, there were no significant differences (p>0.05) in CAT and SOD enzymes activity between paraquat-exposed and control O. niloticus (Figures 2C and 2D, SI 2). Similarly, at day 28, GSH and CAT enzymes activity were not significantly different (p>0.05) between glyphosate-exposed and control O. niloticus (Figures 2F and 2I, SI 2).
However, significant differences (p<0.05) were observed in GSH, GST and MDA activities at day 28 between paraquat-exposed and control O. niloticus (Figures 2A, 2B and 2E, SI 2). Specifically, GSH activity was significantly higher (p<0.05) in 0.11 mg/L concentration compared to 1.12 mg/L concentration of paraquat-exposed and control O. niloticus (Figure 2A, SI 2). Further, GST and MDA activities were significantly higher (p<0.05) in both subacute concentrations of paraquat-exposed O. niloticus when compared to controls (Figure 2B and 2E, SI 2).
Similarly, significant differences (p<0.05) were observed in GST, SOD and MDA activities at day 28 between glyphosate-exposed and control O. niloticus (Figures 2G, 2H and 2J, SI 2). Specifically, GST and MDA activities were significantly higher (p<0.05) (Figures 2G and 2J, SI 2) while SOD activity was significantly lower (p<0.05) at both subacute concentrations of glyphosate-exposed O. niloticus compared to control (Figure 2H, SI 2).
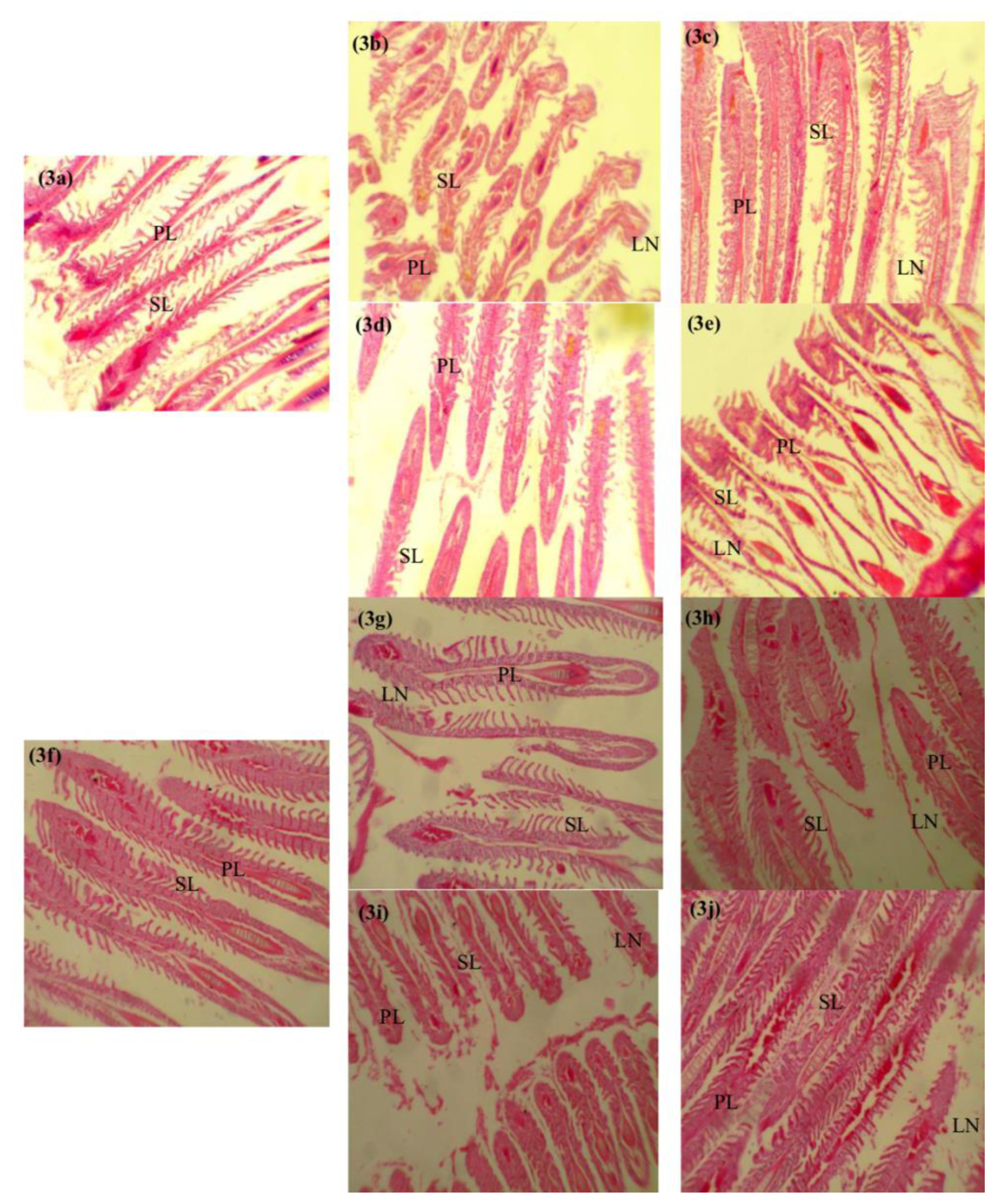
Histological biomarkers in O. niloticus exposed to subacute concentrations of paraquat and glyphosate
Photomicrographs of the representative histological sections from fish in each experimental group is shown in Figures 3 and 4. The histological evaluations revealed mild shortening (0.11 mg/L paraquat, 0.12 mg/L glyphosate) (Figures 3b, 3e, 3g and 3j) to severe shortening (1.12 mg/L paraquat, 0.01 mg/L glyphosate) (Figures 3c, 3h and 3i) of the primary lamella in the gills of exposed O. niloticus respectively at days 14 and 28. Though no abnormalities were observed in the gills of 0.01 mg/L glyphosate exposed O. niloticus at day 14 (Figure 3d) and control O. niloticus at days 14 and 28 (Figures 3a and 3f). Portal inflammatory cells were observed at day 14 in 0.12 mg/L glyphosate-exposed O. niloticus only (Figure 4e) and in all paraquat- and glyphosate-exposed O. niloticus at day 28 (Figures 4g–4j). There were no histological abnormalities in the liver of paraquat-exposed and 0.01 mg/L glyphosate-exposed O. niloticus at day 14 (Figures 4b–4d) and control O. niloticus at days 14 and 28 Figures 4a and 4f.
Discussion
This study evaluated the relative acute toxicity as well as the genotoxic, biochemical and histological biomarkers of exposure to subacute concentrations of the herbicides (paraquat and glyphosate) in O. niloticus (Nile Tilapia). The 96 hLC50 value (11.20 mg/L) of paraquat to O. niloticus in this study is lower than the 96 hLC50 values of 11.84 mg/L and 20 mg/L reported in O. niloticus exposed to paraquat by Babatunde et al. [10] and Figueiredo-Fernandes et al. [11] respectively. Similarly, the 96 hLC50 value (1.22 mg/L) of glyphosate in the current study is lower than the observed LC50 values of 11.30 mg/L [36] and 16.8 ppm [13] in O. niloticus exposed to glyphosate. The observed higher toxicity of glyphosate up to 9x that of paraquat differs with the observations of Ayanda et al. [9] who stated paraquat toxicity up to ~8x that of glyphosate against Clarias gariepinus. Similarly, in an immobility assay with Daphnia magna, paraquat (OSAQUAT) was observed to be ~14x more toxic when compared to glyphosate (RON-DO) based on the 48 hEC50 values [37]. Glyphosate and paraquat have been observed to be moderately toxic and very toxic respectively to aquatic organisms [37,38]. The higher toxicity of glyphosate compared to paraquat in this study may be due to the “extreme” stability of the former in sterile water in the laboratory [37,39]. The differential toxicities of the two herbicides against O. niloticus can be attributed to their different methods of action, fish species, herbicide formulation and life stage [25,40].
A chemical genotoxic agent’s action may result to an increase in micronucleus and nuclear abnormalities occurrence [41]. A vast amount of chemicals can impede the DNA synthesis of an exposed organism and this can bring about nuclear aberrations [42]. The development of these aberrations represents a way to remove from the cell nucleus any elaborated genetic material [43]. The dose-dependent genotoxic biomarkers (nuclear aberrations) observed in the paraquat-exposed fishes especially for MN differs from the observations of Amaeze et al. [44] who observed minimal nuclear abnormalities including no MN at paraquat exposures of 0.1 mg/L (102.84 μg/L). However, the study results corroborate the observations of Oladokun et al. [25] who reported a significant (p<0.05) dose-dependent increase in micronuclei in the red blood cells of the African sharptooth catfish (C. gariepinus) exposed to sublethal concentrations of paraquat at day 28. On the other hand, the observed significant increase in MN in the glyphosate-exposed O. niloticus, though not dose-dependent corroborates the findings of Samantha et al. [17] and Acar et al. [19] who reported nuclear abnormalities in O. niloticus exposed to 17.2 mg/L and different concentrations 5, 10, 20, 30 and 40 mg/L respectively of glyphosate. Several studies have reported pollutants which induce nuclear aberrations in fish tissues [45,46]. Whilst the mechanisms initiating these nuclear aberrations are not fully described, these aberrations are viewed as signs of genotoxic damage, hence, they can match the scoring of these parameters (micronuclei, blebbed nuclei and binucleated cells) during normal genotoxicity surveys [21,47].
Aquatic ecosystems are usually affected by numerous pesticides from various sources [48]. Perturbations at the biochemical and cellular levels are amongst the subtle biological responses observed from exposure of fishes to pollutants [49]. Herbicides can potentially elicit reactive oxygen species (ROS) in living organisms causing oxidative stress in non-target organisms [50]. Defensive mechanisms have emerged from fish and other vertebrates to counter the deleterious ROS effects resulting from various xenobiotics metabolism [51]. CAT and SOD enzymes possess similar roles [52]. SOD is a metalloenzyme that plays an important antioxidant role. It is the major protection countering the toxic superoxide radicals effect in organisms enabling the conversion of superoxide radicals to H2O2 and H2O [53]. Catalase facilitates the removal of H2O2, which metabolizes to molecular oxygen and water [54]. The observed induction of lipid peroxidation (indicated by increased MDA levels) and most antioxidant enzymes in this study especially at day 28 agrees with previous studies reporting alteration in enzyme activities in glyphosate-exposed O. niloticus at concentrations of 1.2 mg/L [15], induction of hepatic oxidative stress at 5, 10, 20, 30 and 40 mg/L [19] and reduction in antioxidant enzymes activity at 0.2, 0.8, 4 and 16 mg/L [18] of glyphosate. Further, this study revealed a significant decrease in SOD level at day 28 following exposure O. niloticus to subacute concentrations of glyphosate. This could be a reaction to the elevated ROS induced by the toxic effects of herbicides [55]. The observed significantly increased GST levels in the herbicides concentrations and significantly increased GSH level in the lower paraquat concentration at day 28 only contrasts with the findings of Moustafa et al. [56] who observed a significant reduction in GSH and GST levels in C. gariepinus exposed to glyphosate. The major function of GSH is to protect cells against oxidative stress due to free radicals through reductive detoxification of reactive intermediates like H2O2 [57]. On the other hand, GSTs potentiates the coupling of reduced GSH with electrophilic metabolites. Also, they are involved in depuration of reactive intermediates and oxygen radicals [54]. Studies have shown that activities of these enzymes may be potentiated in the liver of fish exposed to various pollutants [50].
Histology studies are regarded as a sensitive endpoint in detecting organ toxicity to xenobiotics [58] with capabilities of revealing detailed information regarding the acute and chronic effects of toxicants on targeted organs which may not be detected by functional biomarkers [59]. Fish gills are the main place for ion exchange with the environment, and also, the main channel of pesticide penetration, this is because they are in constant contact with water [60]. The histological alterations observed in this study ranging from mild to severe lamellae necrosis especially at day 28 in the herbicides-exposed fishes agrees with the observations of liver and gonadal pathologies as well as gill pathologies in O. niloticus exposed to paraquat at 0.5 mg/L [11] as well as 12 mg/L and 14.20 mg/L [10] respectively. Similarly, the study findings agree with previous studies observations in glyphosate-exposed O. niloticus of gill, liver and kidney pathologies at 5, 12 and 16.8 ppm [13,14] as well as at 17.2 mg/L [16]. Several studies have revealed alterations in O. niloticus liver elicited by different toxic chemicals [61]. Fusion of secondary lamellae as a result of exposure to pesticides appears to have a protective role in diminishing the affected gill surface; this response slows down the penetration of toxic and may result in fish choking [62, 63].
Conclusions
This study revealed significant biomarkers of subacute concentrations of paraquat and glyphosate in O. niloticus. The novelty of this study is the comparative assessment of multiple biomarkers at varying levels of biological organization (from cell to whole system level) of two (2) commonly used herbicides (paraquat and glyphosate) at subacute concentrations (near environmentally relevant concentrations). The observed toxicities may pose ecological risk through potential bioaccumulation of the herbicides in non-target aquatic organisms and biomagnification through the food chain. Hence, studies and advocacies on risk of herbicides use with potential attendant impact on non-target organisms are essential [64]. Appropriate and irregular herbicides use are advised so that the advantageous effects of these chemicals will be achieved. These will promote responsible consumption and production and sustain life below water (United Nations Sustainable Development Goals 12 and 14 respectively).