Introduction
The gradual loss of structure or function of neurons, that also involves neuronal death often result in diseases or conditions that affect mental functioning and proper movement [1]. Environmental factors are part of the contributors to the pathogenesis of neurodegenerative diseases [2]. Lead (Pb) is one of the well-known environmental toxins that cause neurodegeneration, affect neurodevelopment and mental health [3–6].
Pb is a heavy metal that is widely used in various forms despite well-documented reports of its toxic effects [7]. Exposure to Pb, even in low doses, results in various adverse consequences, particularly in the brain [8]. Pb, as a neurotoxicant, crosses the blood-brain barrier to cause oxidative stress [9], morphologic damage, neurodegeneration and cognitive impairment in the brain, particularly that of developing brain [10–13]. The prefrontal cortex (PFC), which accounts for one-third of the human cerebral cortex, is responsible for working memory, higher reasoning, decision making and many other executive functions [14, 15]. There are evidences that intrinsic connections of the dorsolateral PFC (DLPFC) (Brodmann areas 9 & 46) make more connections with other parts of PFC which are relevant to cognitive processing [16–18].
Since neurotoxicity by Pb was confirmed as one of the causative agents of neurodegeneration, emerging evidence suggests that dietary phytochemicals have potentials to chelate its effects. In this sense, flavonoid may act on several therapeutic targets to prevent or reduce the events involved in neurodegeneration caused by Pb neurotoxicity,
Flavonoids have been proposed to limit neurodegeneration in diverse neurological diseases, including cognitive impairments [19, 20]. According to emerging evidence, dietary flavonoids and their metabolites cross the blood-brain barrier and exert multiple neuropharmacological actions which include modulating the innate architecture of the brain to impact cognitive function and have a neuroprotective effect [19]. Crucial factors that are involved in inducing synaptic plasticity, suppression of neuroinflammatory processes, promotion of cerebrovascular perfusion, and activation of adaptive cellular stress response to protect against oxidative stressors by flavonoids are regulation of neuronal signalling cascades and transcription factors [19, 20]. Vitexin is one of flavonoids that is rapidly gaining the attention of researchers because of is various potentials.
Vitexin is a flavonoid that is extracted from various plants including pearl millet, hawthorn, pigeon pea, mung bean, mosses, Passiflora, bamboo, mimosa, wheat leaves, and chaste berry [21]. Its effects are reported in pharmacological roles range from anti-oxidative anti tumour, anti-viral, anti-inflammatory, anti-bacterial, antihypertensive, antinociceptive, antispasmodic, anti-diabetic, antidepressant, neuroprotective and cognitive improving functions [22–28]. The contribution of Vitexin to neuroprotection is through counteracting the targets that induce neurodegeneration, such as redox imbalance, neuroinflammation and abnormal protein aggregation, which then lead to a reduction of cognitive or motor impairment [24].
Although the preventive effects of Vitexin on neurodegeneration could be considered mostly substantiated, the positive neuroprotective activity after acute Pb administration still deserves more research. Hence, the aim of the study was to investigate the neuroprotective effect of Vitexin against Pb induced neurodegenerative changes in the DLPFC and working memory impairment in mice.
Materials and methods
Treatment solution
Vitexin extract and Pb acetate were purchased from Sigma-Aldrich (St Louis, MO, USA). Phosphate buffered saline was freshly prepared, while other materials were purchased locally. Pb acetate solution was prepared by dissolving 500 mg of Pb in 1 mL water. Vitexin solution was freshly prepared each morning of administration before use. Vitexin was dissolved in normal saline (NaCl, 0.9%). Pb treatments on mice were done using oral gavage, while all Vitexin treatment was via intraperitoneal injection (IP).
Animals
Thirty-two adolescent male albino mice (weight: 17–18g) were procured from the Temidale animal husbandry centre, Ogbomosho, Nigeria. The animals were acclimatized for 14 days on a 12 h of light/dark cycle at the animal holdings of Central Research Lab, of the University of Ilorin, Nigeria, before administration. The animals were maintained under the National Institutes of Health guide for the care and use of laboratory animals (NIH Publications, 8th edition). The University of Ilorin Ethical and Review Committee approved the experimental protocols used in the study (number: UERC/ASN/2018/1257).
Animal grouping and administration
The animals were randomly divided into four groups of eight animals. Group A which was also the control group was administered 0.2 mL normal saline for 14 days via oral gavage; Group B (Pb group) received oral gavage of Pb acetate 100 mg/kgbw-d for 14 days; Group C (Vitexin group) received intraperitoneal injection of Vitexin 1 mg/kgbw-d for 14 days; Group D (Pb+Vitexin group) received concurrent administration of 100 mg/kgbw-d of Pb + 1 mg/kgbw-d Vitexin for 14 days.
Barnes maze test
This test for spatial learning memory was done between 21st to 25th day after the end of administration according to the modified methods in literature [29]. In this test, a circular table with 15 holes around the circumference, was placed in a lighted room. Fourteen holes of them lead to an open drop to the floor, but a single hole leads to a dark box that the animal can hide. In initial trials, the animals were placed in the centre of the table and were led to the dark box. In subsequent trials, the animals were placed in the centre of the table and were to find the dark box on their own. The test was recorded using a video camera attached to a computer. The record of the number of incorrect holes visited before the correct hole was also taken. The time it took for the animals to find the correct hole was recorded.
Novel object recognition (NOR) test
This test was carried out between 26th to 30th day after administration according to the method described in previous literature [30]. The first two days of this test was for habituation; the third was the trial while the fourth day was the test proper. At the end of this test, the percentage preference for the new object was calculated,
Preparation of brain sample
At the end of the last neurobehavioral test, the animals were anaesthetized with an intraperitoneal injection of ketamine (90 mg/kg). The mice were transcardially perfused with 4% paraformaldehyde (PFA) in phosphate buffer saline (PBS) solution. Excised PFC tissues were postfixed in 4% PFA for 24 hours before transferring into a cold 20% sucrose solution. Tissue was processed as paraffin wax embedded blocks, while haematoxylin and eosin (H&E), cresyl fast violet (CFV) and congo red staining procedures, were carried out [31].
Photomicrography and cell count
Stained sections were viewed under a light binocular microscope (Olympus, NJ, USA) attached to an amscope camera (MD500, CA, USA). Six visual fields of CFV stain of the DLPFC were photographed in each section and were used for cell counting as well as other morphometric analysis with the aid of ImageJ 1.52r software (National Institute of Health, USA). The DLPFC was identified [32].
Statistical analysis
All statistical analyses and graph fitting were performed using Graph Pad Prism version 5.0 for Windows. The significance of difference was determined using one-way ANOVA followed by Tukey’s posthoc test for all parameters. All data were expressed as the mean ± standard error of the mean (SEM), with n representing the number of animals used in each experiment. Statistical significance was defined at the level of p < 0.05.
Results
Effect of Vitexin and lead (Pb) acetate on learning
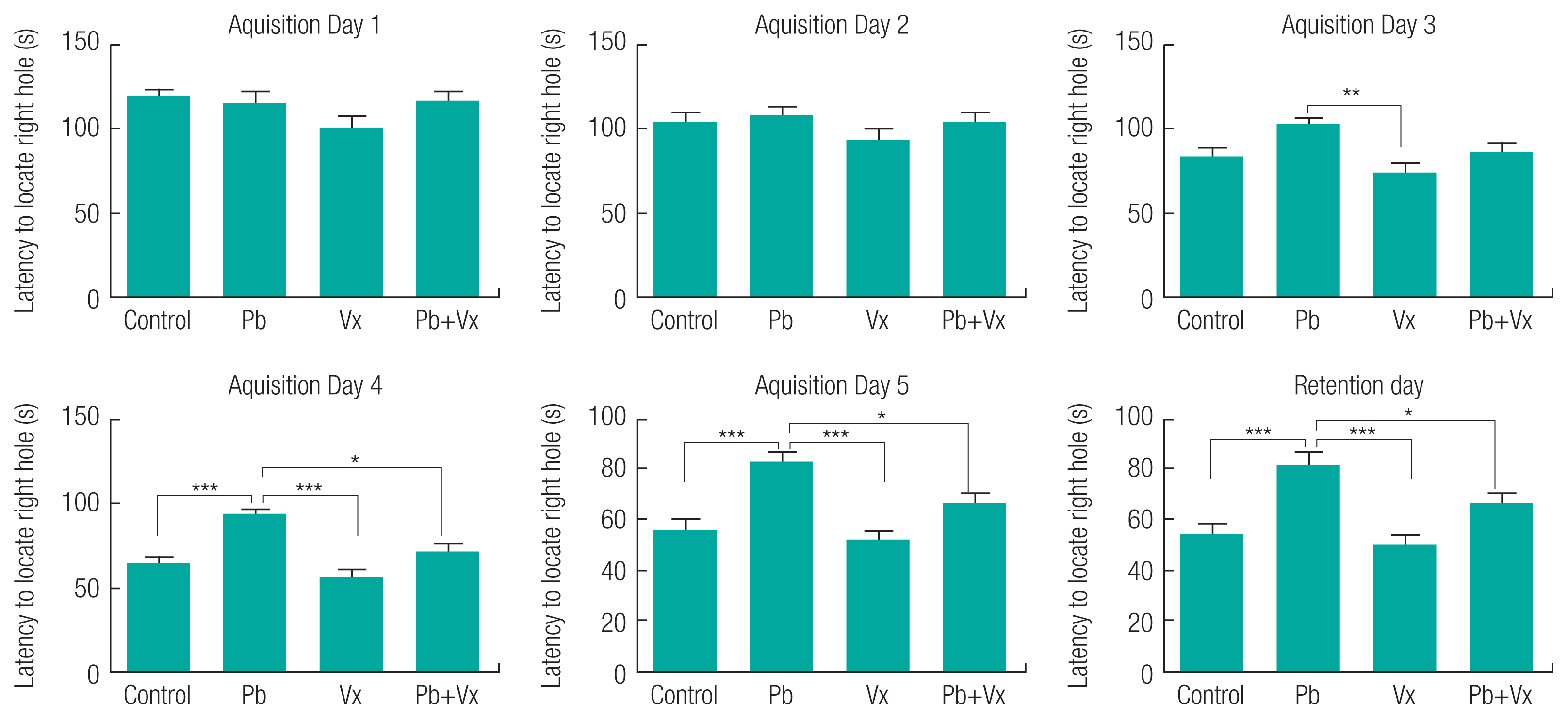
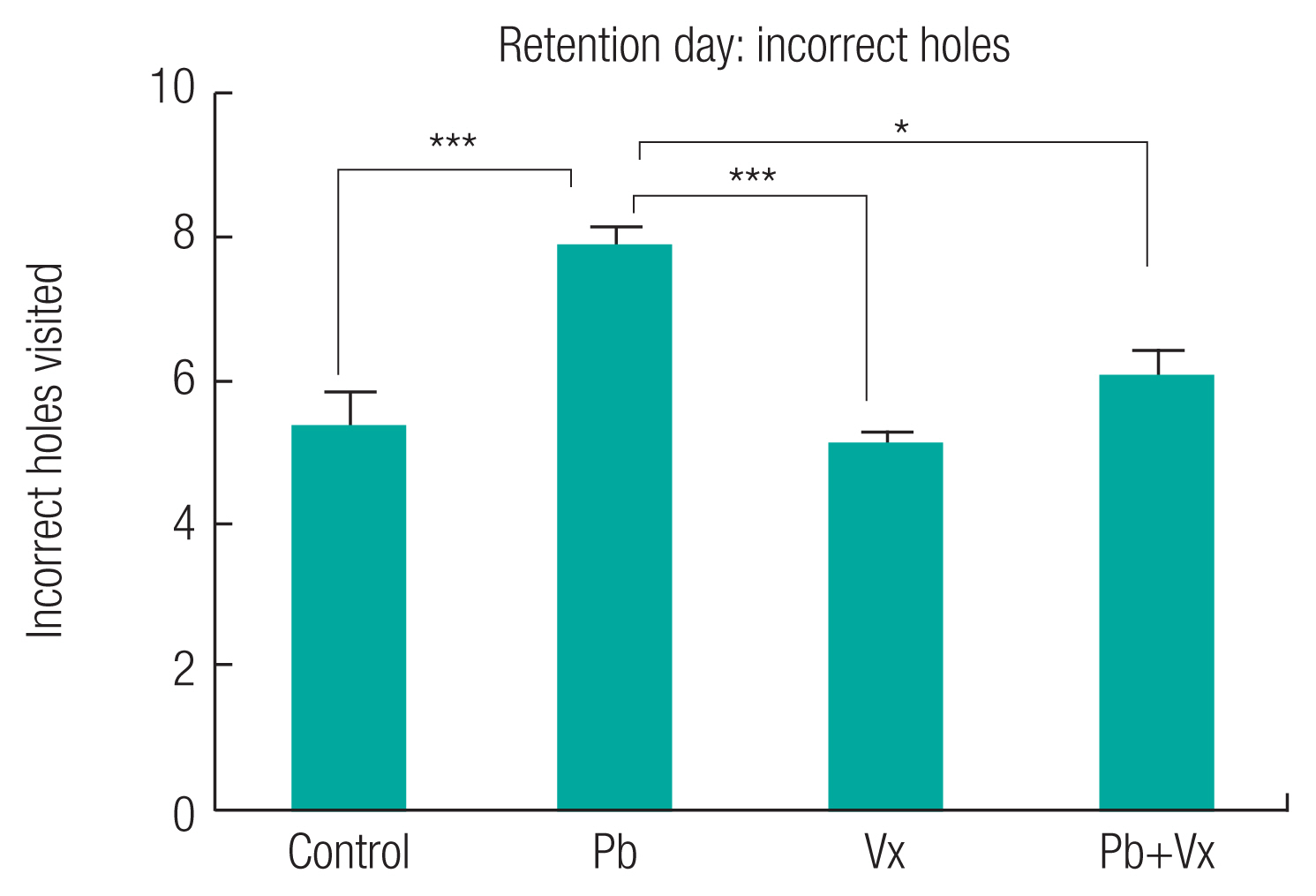
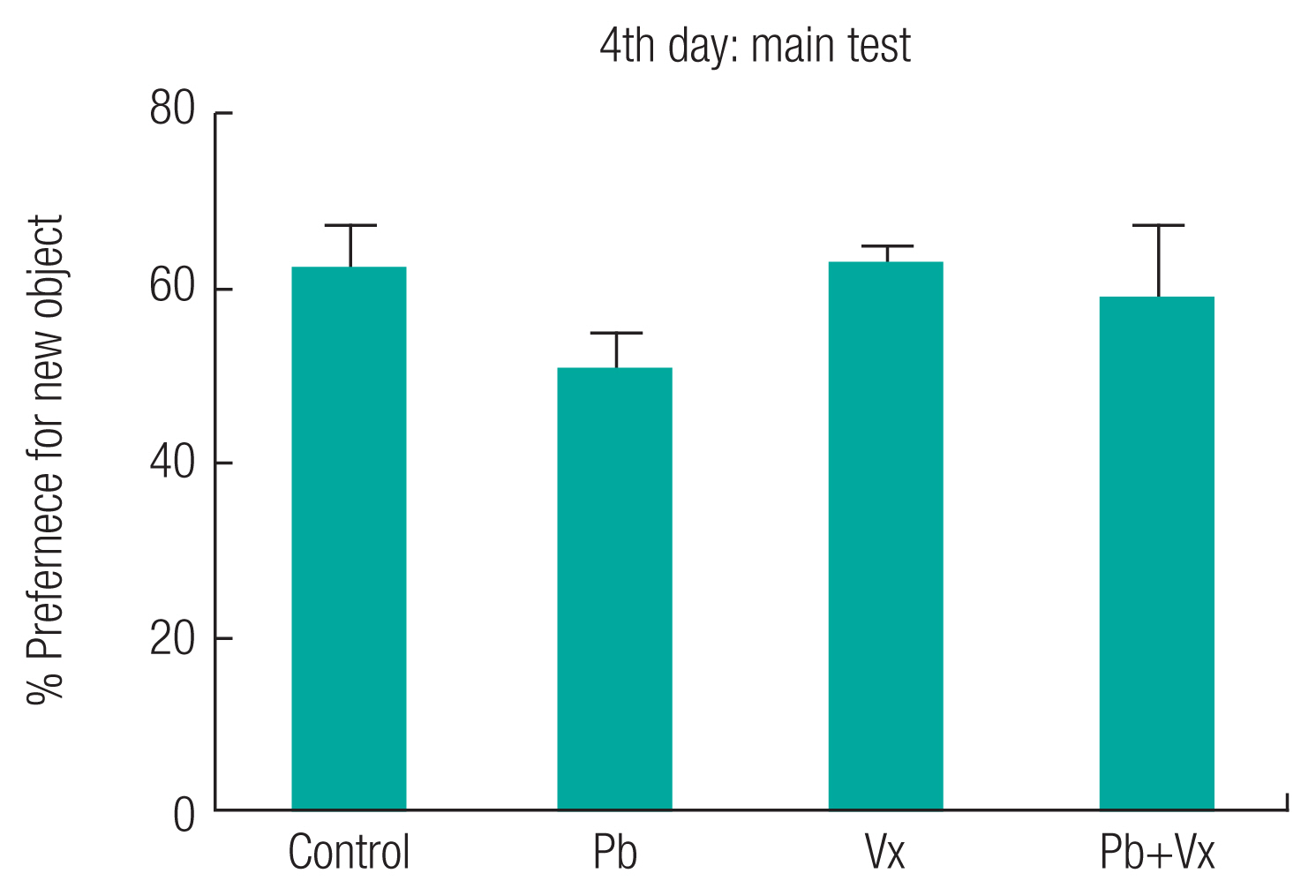
The result from the Barnes maze test (Figure 1) shows no statistically significant difference (p > 0.05) when the Pb group was compared with the other groups within the first three days of learning acquisition. Starting from the fourth day (retention day), the time it took animals to locate the dark box in control, vitexin, and Pb+Vitexin groups became significantly shorter when compared with the Pb group (p <0.001; p <0.001 and p <0.05 respectively). Similarly, the number of incorrect holes visited by the animals before finding the dark box on the retention day (Figure 2) was significantly reduced in the control, Vitexin, and Pb+Vitexin groups (p <0.001; p <0.001 and p <0.05 respectively) when comparisons were made with Pb group. Furthermore, in the NOR test, the percentage preference of the animals (Figure 3) to a new object, was generally higher in control, Vitexin, and Pb+Vitexin groups than the Pb group. However, the difference in percentage preference of the animals to a new object was not statistically significant.
Histoarchitectural observation
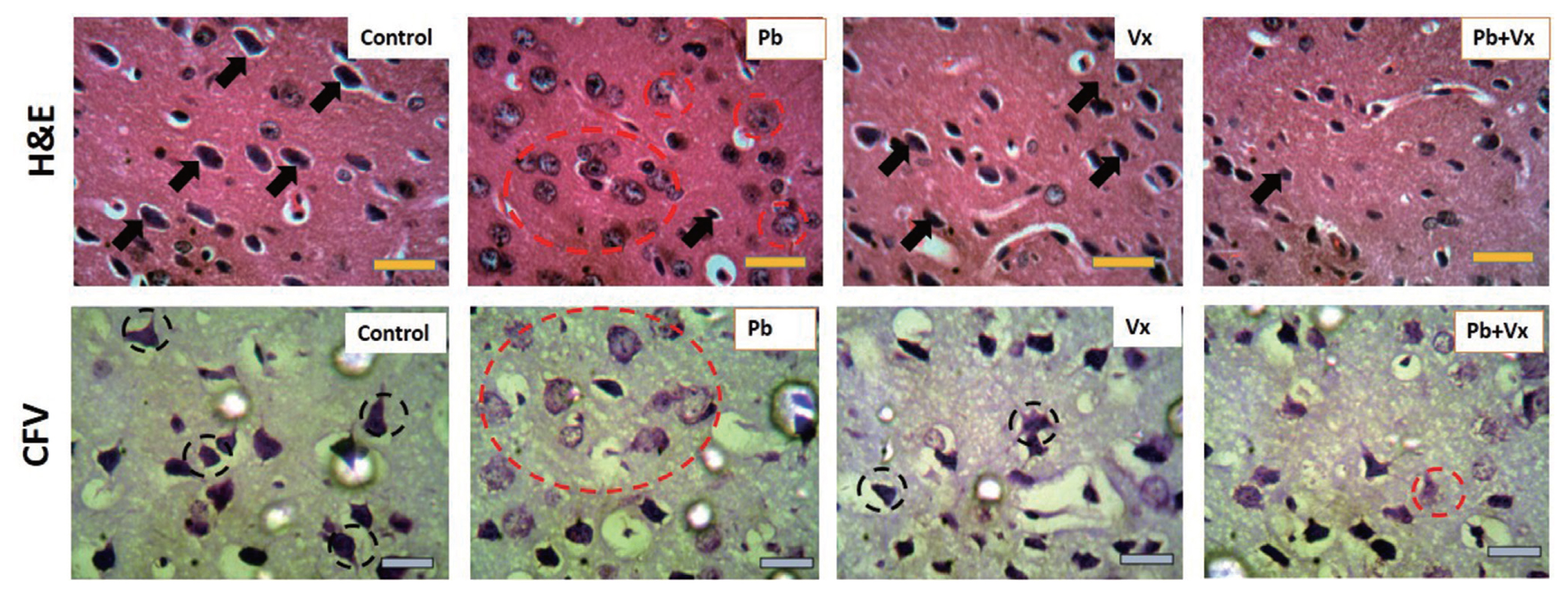
The representative photomicrograph of H&E-stained sections of mice DLPFC at higher magnification showed the control group with normal cytoarchitecture that was characterized by large pyramidal neurons (black arrows in Figure 4) with small perineural spaces. The Pb group showed degenerative changes that were characterized by numerous pyknotic pyramidal neurons that appear with fragmented cytoplasm and condensed nuclei within the soma (dotted red circles in Figure 4). Vitexin group present similar cytoarchitecture to the control group. The Pb+Vitexin group showed recovery in the histoarchitecture when compared to the degenerative changes that were observed in the Pb group.
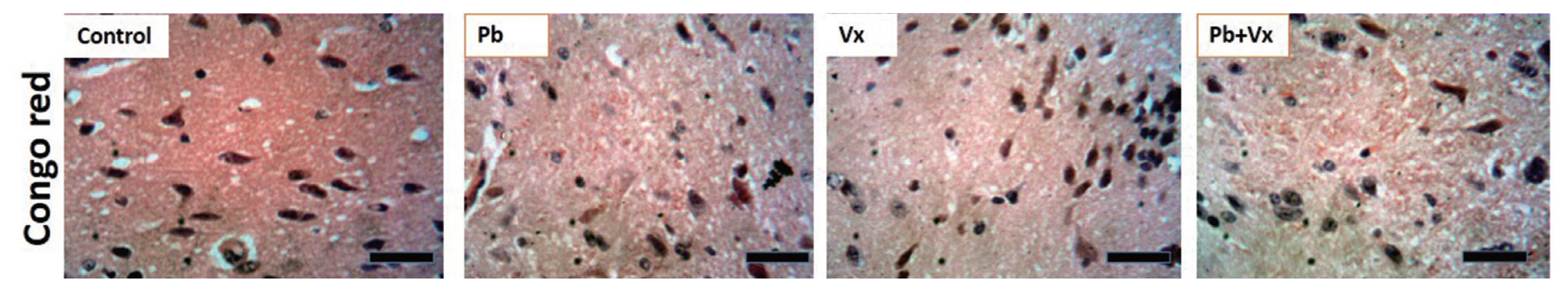
The representative photomicrograph of CFV stained sections of mice DLPFC showed the control and Vitexin groups stain intensely for Nissl (black dotted circle in Figure 4) with very few chromatolytic cells. However, in Pb group, numerous cells undergo chromatolysis (red dotted circle in Figure 4). In the Pb+Vitexin group, there were fewer chromatolytic cells when compared to the observation in Pb group. Furthermore, there appear to be no amyloid aggregation in any of the groups when the sections were stained with congo red (Figure 5).
In Figure 6, it was observed that there were more degenerating neurons in Pb group than healthy neurons when the count was analyzed (p <0.001). The result was contrary to the observation in others where healthy neurons were significantly higher than degenerating neurons. In control, Vitexin, and Pb+Vitexin groups, the p values between normal and degenerating neuron count were all <0.001.
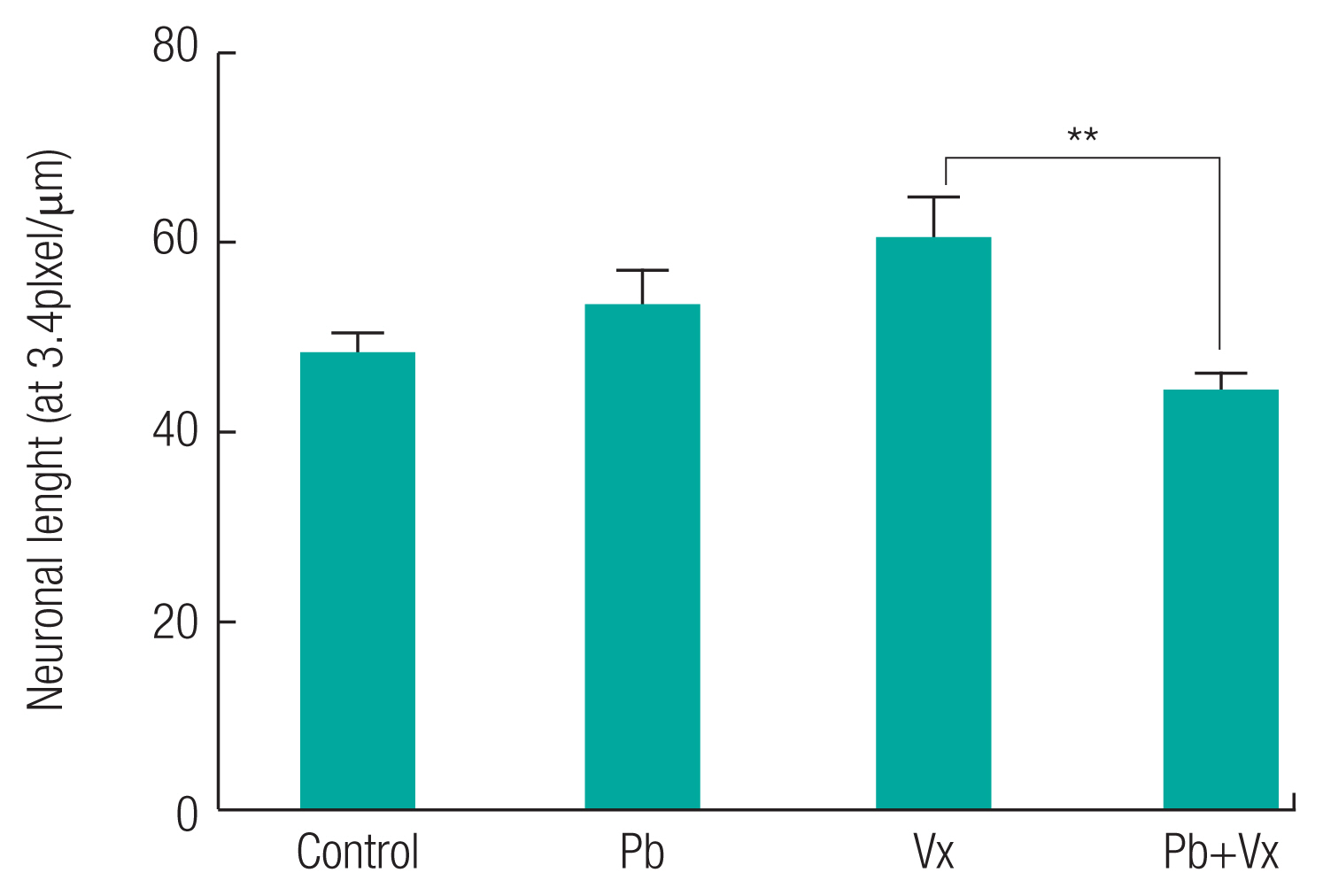
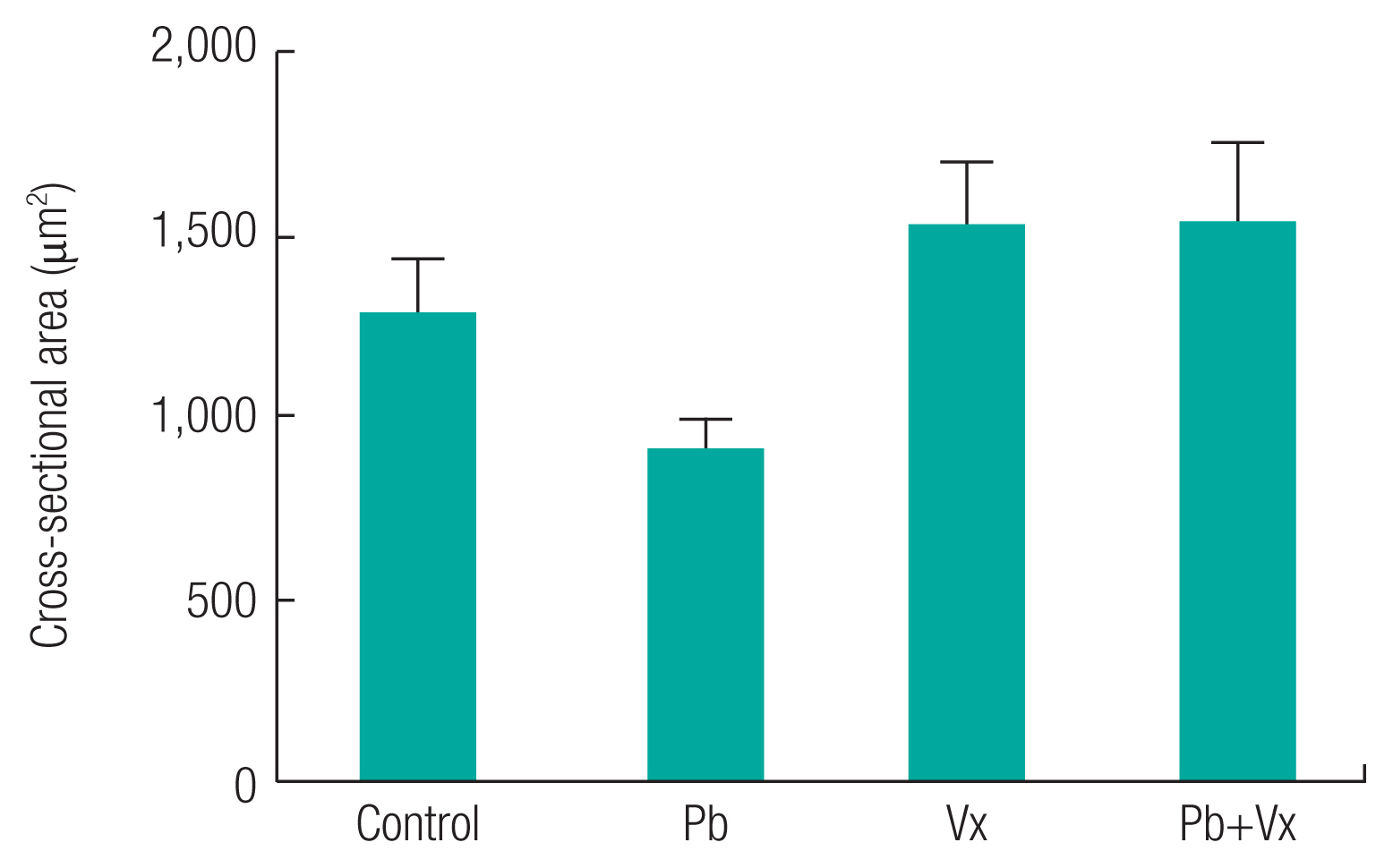
Intercellular distance between healthy neurons in each of the groups were not statistically significant (p >0.05) when comparisons were made between the groups (Figure 7). In addition to the result seen, the neuronal length in control, Vitexin, and Pb+Vitexin groups was not statistically different (p >0.05) when compared with the Pb group. However, neuronal length in Pb+Vitexin group was significantly smaller (p <0.01) when compared with the Vitexin group. The cross-sectional area of neurons (Figure 8) were not statistically different (p >0.05) between the groups but still appear lowest in Pb group when compared with either Vitexin or Pb+Vitexin groups.
In Table 1, the thickness of layer II–III of DLPFC was significantly small in Pb group when compared with that in the control, Vitexin, and Pb+Vitexin group. Similarly, the thickness of layer II–III of DLPFC in the Pb+Vitexin group was significantly small when compared with that in either the control or the Vitexin group.
Discussion
This study shows that animals in Pb-treated group took longer time to locate the dark box when compared with the group that had concurrent treatment of Pb+Vitexin (Figure 1). The result also shows that animals in Pb-treated group visited more incorrect holes before locating the dark box (Figure 2). This result indicates that Pb treatment may have affected the ability of the animals to learn or remember the location of the dark box earlier than expected after several trial period. This finding supported several earlier reports that implicated Pb exposure as an agent of cognitive impairment, particularly in a developing brain [13, 33–35]. This result further indicates that the treatment of Vitexin alongside Pb treatment will attenuate the possible cognitive impairment caused by Pb. Earlier reports have indicated the various roles of Vitexin to improve cognitive impairment that was not caused by Pb exposure [36, 37]. According to Moosavirad et al., Pb-exposure cause reduction in the discrimination and object recognition index of learning tasks [38]. However, in this study, Pb treated group did not significantly show a reduction in preference for a new object when compared with Pb+Vitexin group (Figure 3). The differences observed could be as a result of differences in the duration of administration between the two studies.
The histoarchitectural observations in this study, shows that in groups that were treated with Pb only, degenerative changes that were characterized by numerous pyknotic neurons with fragmented cytoplasm and condensed nuclei were observed (Figure 4). Also lightly stained Nissl and numerous chromatolytic cells were observed. Several earlier studies had reported the ability of Pb to cause neuronal degeneration, chromatolysis and distortion of cortical layers [39–42]. This result indicates that the number of healthy neurons available to carry out normal function within the DLPFC will be low, thereby affecting is the function in working memory (Figure 1). In the group, where both Pb and Vitexin was concurrently administered, there was a significant improvement in a possible degenerative change and chromatolysis was significantly reduced (Figure 4). As part of previous studies, Vitexin has demonstrated its ability to decrease neuronal cell death [43, 44] through a different mechanism and to significantly improve neuronal cell structure and function [24, 45]. This result in the Pb+Vitexin group, indicates that the number of healthy neurons in DLPFC will be more than the degenerated ones when compared to Pb group (as Figure 4, 6). In this study, amyloid aggregation was not observed in any of the groups probably because of the dose and duration of treatment used or the type of microscope used to capture the stained sections (Figure 5).
This study shows that there was no significant difference in intercellular distance, neuronal length, and the cross-sectional area of neurons in Pb group when compared with the Pb+Vitexin group. This result indicates that regardless of Pb only treatment or Pb+Vitexin treatment, the intercellular distance, neuronal length and the cross-sectional area of neurons remain the same. Furthermore, the thickness of layer II–III of DLPFC was significantly small when compared with the Pb+Vitexin group. According to an earlier report by Defelipe et al., layer 2/3 of mouse cortical thickness was 1210 μm [46]. The result of this study indicates that the number of components of layer II–III of DLPFC in Pb group would be smaller than Pb+Vitexin group.
Conclusion
The result of this study has shown that concurrent administration of Vitexin and Pb will significantly reduce neurodegeneration and improve working memory. However, Pb treatment or both Pb and Vitexin treatment does not have an effect on intercellular distance, neuronal length and cross-sectional area of neurons in layer III of DLPFC.